

Centromeric chromatin pliability and memory at a human neocentromere
- PMID: 12743043
- PMCID: PMC155990
- DOI: 10.1093/emboj/cdg232
Centromeric chromatin pliability and memory at a human neocentromere
Abstract
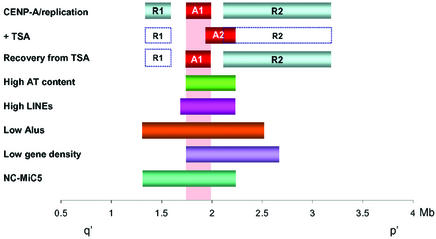
We show that Trichostatin A (TSA)-induced partial histone hyperacetylation causes a unidirectional shift in the position of a previously defined binding domain for the centromere-specific histone H3 homologue CENP-A at a human neocentromere. The shift of approximately 320 kb is fully reversible when TSA is removed, but is accompanied by an apparent reduction in the density of CENP-A per unit length of genomic DNA at the neocentromere. TSA treatment also instigates a reversible abolition of a previously defined major domain of differentially delayed replication timing that was originally established at the neocentromeric site. None of these changes has any measurable deleterious effects on mitosis or neocentromere function. The data suggest pliability of centromeric chromatin in response to epigenetic triggers, and the non-essential nature of the regions of delayed replication for centromere function. Reversibility of the CENP-A-binding position and the predominant region of delayed replication timing following removal of TSA suggest strong memory at the original site of neocentromeric chromatin formation.
Figures
References
-
- Aagaard L., Schmid,M., Warburton,P. and Jenuwein,T. (2000) Mitotic phosphorylation of SUV39H1, a novel component of active centromeres, coincides with transient accumulation at mammalian centromeres. J. Cell Sci., 113, 817–829. - PubMed
-
- Barry A.E., Howman,E.V., Cancilla,M.R., Saffery,R. and Choo,K.H.A. (1999) Sequence analysis of an 80 kb human neocentromere. Hum. Mol. Genet., 8, 217–227. - PubMed
-
- Bickmore W.A. and Carothers,A.D. (1995) Factors affecting the timing and imprinting of replication on a mammalian chromosome. J. Cell Sci., 108, 2801–2809. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
