

The herpes simplex virus 1 US3 protein kinase blocks caspase-dependent double cleavage and activation of the proapoptotic protein BAD
- PMID: 12743316
- PMCID: PMC155029
- DOI: 10.1128/jvi.77.11.6567-6573.2003
The herpes simplex virus 1 US3 protein kinase blocks caspase-dependent double cleavage and activation of the proapoptotic protein BAD
Abstract
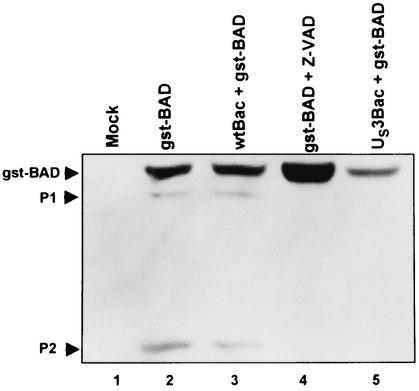
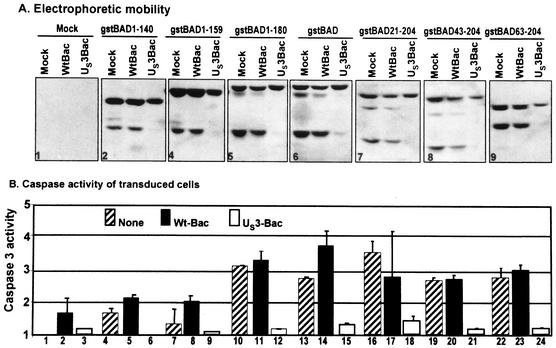
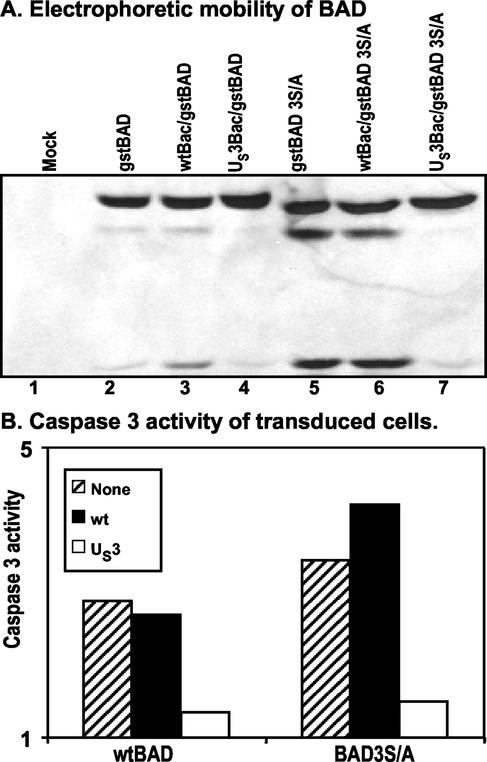
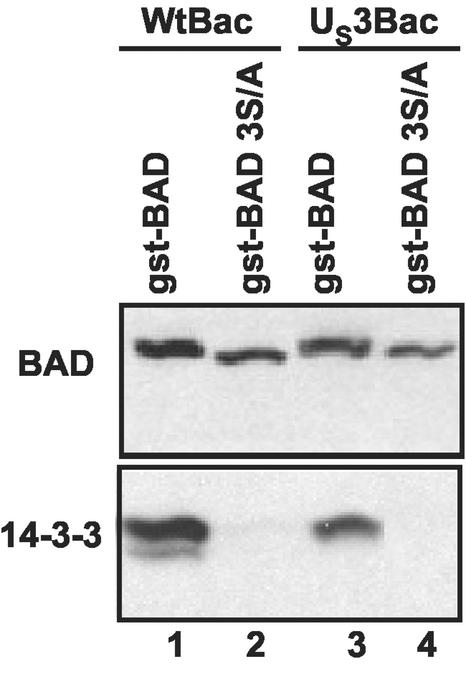
An earlier report showed that the U(S)3 protein kinase blocked the apoptosis induced by the herpes simplex virus 1 (HSV-1) d120 mutant at a premitochondrial stage. Further studies revealed that the kinase also blocks programmed cell death induced by the proapoptotic protein BAD. Here we report the effects of the U(S)3 protein kinase on the function and state of a murine BAD protein. Specifically, (i) in uninfected cells, BAD was processed by at least two proteolytic cleavages that were blocked by a general caspase inhibitor. The untreated transduced cells expressed elevated caspase 3 activity. (ii) In cells cotransduced with the U(S)3 protein kinase, the BAD protein was not cleaved and the caspase 3 activity was not elevated. (iii) Inasmuch as the U(S)3 protein kinase blocked the proapoptotic activity and cleavage of a mutant (BAD3S/A) in which the codons for the regulatory serines at positions 112, 136, and 155 were each replaced with alanine codons, the U(S)3 protein kinase does not act by phosphorylation of these sites nor was the phosphorylation of these sites required for the antiapoptotic function of the U(S)3 protein kinase. (iv) The U(S)3 protein kinase did not enable the binding of the BAD3S/A mutant to the antiapoptotic proteins 14-3-3. Finally, (v) whereas cleavage of BAD at ASP56 and ASP61 has been reported and results in the generation of a more effective proapoptotic protein with an M(r) of 15,000, in this report we also show the existence of a second caspase-dependent cleavage site most likely at the ASP156 that is predicted to inactivate the proapoptotic activity of BAD. We conclude that the primary effect of U(S)3 was to block the caspases that cleave BAD at either residue 56 or 61 predicted to render the protein more proapoptotic or at residue 156, which would inactivate the protein.
Figures

References
-
- Adachi, M., and K. Imai. 2002. The proapoptotic BH3-only protein BAD transduces cell death signals independently of its interaction with Bcl-2. Cell Death Differ. 9:1240-1247. - PubMed
-
- Asano, S., T. Honda, F. Goshima, D. Watanabe, Y. Miyake, Y. Sugiura, and Y. Nishiyama. 1999. US3 protein kinase of herpes simplex virus type 2 plays a role in protecting cornel epithelial cells from apoptosis in infected mice. J. Gen. Virol. 80:51-56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
