

Progesterone and glucocorticoid receptors recruit distinct coactivator complexes and promote distinct patterns of local chromatin modification
- PMID: 12748280
- PMCID: PMC155204
- DOI: 10.1128/MCB.23.11.3763-3773.2003
Progesterone and glucocorticoid receptors recruit distinct coactivator complexes and promote distinct patterns of local chromatin modification
Abstract
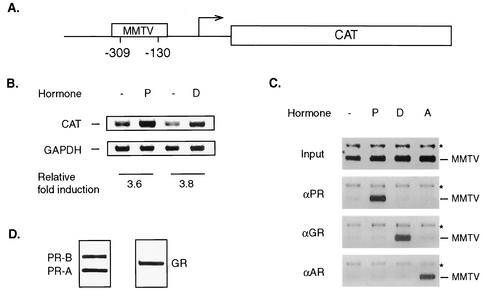
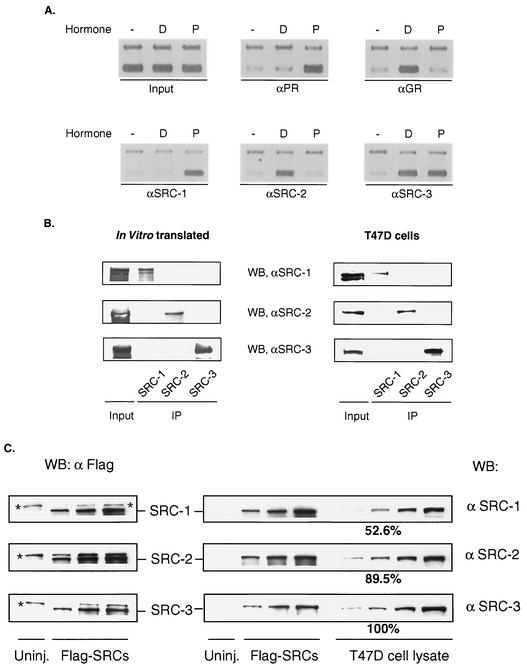
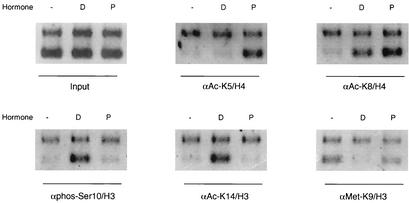
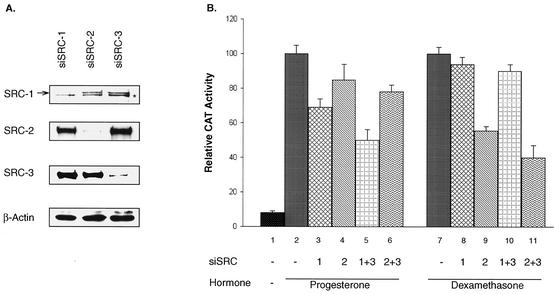
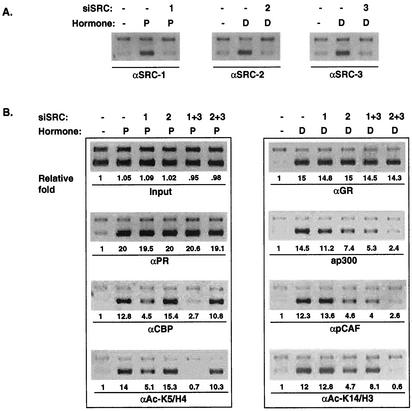
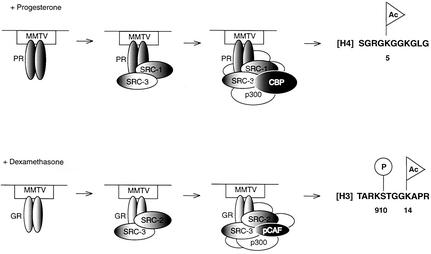
It is well established that steroid receptor function requires interaction with coactivators. However, the mechanisms through which steroid receptors elicit precise assembly of coactivator complexes and the way the steroid activation signal is transduced remain elusive. Using a T47D cell line stably integrated with a mouse mammary tumor virus-chloramphenicol acetyltransferase (MMTV-CAT) reporter, we demonstrate that specific steroid receptors exhibit preferential recruitment of SRC-1 family coactivators, which determines the subsequent recruitment of specific downstream coregulator molecules. Upon ligand treatment, progesterone receptor (PR) interacted preferentially with SRC-1, which recruited CBP and significantly enhanced acetylation at K5 of histone H4. In contrast, activated glucocorticoid receptor (GR) preferentially associated with SRC-2 (TIF-2/GRIP-1), which subsequently recruited pCAF and led to specific modification of histone H3, suggesting that specific coactivators recruit distinct histone acetyltransferases to modulate the transcription of steroid-responsive genes. Loss-of-function experiments further support the predicted roles of SRC-1 and SRC-2 in, respectively, PR- and GR-mediated transcription on the MMTV promoter. This study indicates that differential recruitment of coactivators by nuclear receptors determines the assembly of coactivator complexes on target promoters to mediate specific transcription signals.
Figures

Similar articles
-
Mediator and p300/CBP-steroid receptor coactivator complexes have distinct roles, but function synergistically, during estrogen receptor alpha-dependent transcription with chromatin templates.Mol Cell Biol. 2003 Jan;23(1):335-48. doi: 10.1128/MCB.23.1.335-348.2003. Mol Cell Biol. 2003. PMID: 12482985 Free PMC article.
-
E1A-mediated repression of progesterone receptor-dependent transactivation involves inhibition of the assembly of a multisubunit coactivation complex.Mol Cell Biol. 2000 Mar;20(6):2138-46. doi: 10.1128/MCB.20.6.2138-2146.2000. Mol Cell Biol. 2000. PMID: 10688660 Free PMC article.
-
CBP recruitment and histone acetylation in differential gene induction by glucocorticoids and progestins.Mol Endocrinol. 2003 Jun;17(6):1085-94. doi: 10.1210/me.2001-0183. Epub 2003 Mar 13. Mol Endocrinol. 2003. PMID: 12637584
-
Steroid hormone receptor status defines the MMTV promoter chromatin structure in vivo.J Steroid Biochem Mol Biol. 1995 Jun;53(1-6):421-9. doi: 10.1016/0960-0760(95)00088-h. J Steroid Biochem Mol Biol. 1995. PMID: 7626491 Review.
-
Nuclear receptors: coactivators, corepressors and chromatin remodeling in the control of transcription.J Mol Endocrinol. 1999 Dec;23(3):255-75. doi: 10.1677/jme.0.0230255. J Mol Endocrinol. 1999. PMID: 10601972 Review.
Cited by
-
Human papillomavirus E7 oncoprotein dysregulates steroid receptor coactivator 1 localization and function.J Virol. 2006 Jul;80(13):6669-77. doi: 10.1128/JVI.02497-05. J Virol. 2006. PMID: 16775354 Free PMC article.
-
Estradiol downregulation of the tumor suppressor gene BTG2 requires estrogen receptor-alpha and the REA corepressor.Int J Cancer. 2009 Apr 15;124(8):1841-51. doi: 10.1002/ijc.24133. Int J Cancer. 2009. PMID: 19117054 Free PMC article.
-
Androgen-induced activation of gonadotropin-regulated testicular RNA helicase (GRTH/Ddx25) transcription: essential role of a nonclassical androgen response element half-site.Mol Cell Biol. 2012 Apr;32(8):1566-80. doi: 10.1128/MCB.06002-11. Epub 2012 Feb 13. Mol Cell Biol. 2012. PMID: 22331472 Free PMC article.
-
Structure and function of steroid receptor AF1 transactivation domains: induction of active conformations.Biochem J. 2005 Nov 1;391(Pt 3):449-64. doi: 10.1042/BJ20050872. Biochem J. 2005. PMID: 16238547 Free PMC article. Review.
-
Metabolic syndrome without obesity: Hepatic overexpression of 11beta-hydroxysteroid dehydrogenase type 1 in transgenic mice.Proc Natl Acad Sci U S A. 2004 May 4;101(18):7088-93. doi: 10.1073/pnas.0305524101. Epub 2004 Apr 26. Proc Natl Acad Sci U S A. 2004. PMID: 15118095 Free PMC article.
References
-
- Alen, P., F. Claessens, E. Schoenmakers, J. V. Swinnen, G. Verhoeven, W. Rombauts, and B. Peeters. 1999. Interaction of the putative androgen receptor-specific coactivator ARA70/ELE1alpha with multiple steroid receptors and identification of an internally deleted ELE1beta isoform. Mol. Endocrinol. 13:117-128. - PubMed
-
- Allan, G. F., N. H. Ing, S. Y. Tsai, G. Srinivasan, N. L. Weigel, E. B. Thompson, M. J. Tsai, and B. W. O'Malley. 1991. Synergism between steroid response and promoter elements during cell-free transcription. J. Biol. Chem. 266:5905-5910. - PubMed
-
- Bhakat, K. K., and S. Mitra. 2000. Regulation of the human O(6)-methylguanine-DNA methyltransferase gene by transcriptional coactivators cAMP response element-binding protein-binding protein and p300. J. Biol. Chem. 275:34197-34204. - PubMed
-
- Borud, B., T. Hoang, M. Bakke, A. L. Jacob, J. Lund, and G. Mellgren. 2002. The nuclear receptor coactivators p300/CBP/cointegrator-associated protein (p/CIP) and transcription intermediary factor 2 (TIF2) differentially regulate PKA-stimulated transcriptional activity of steroidogenic factor 1. Mol. Endocrinol. 16:757-773. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous