

Short- and long-term enhancement of excitatory transmission in the spinal cord dorsal horn by nicotinic acetylcholine receptors
- PMID: 12748382
- PMCID: PMC164528
- DOI: 10.1073/pnas.1131709100
Short- and long-term enhancement of excitatory transmission in the spinal cord dorsal horn by nicotinic acetylcholine receptors
Abstract
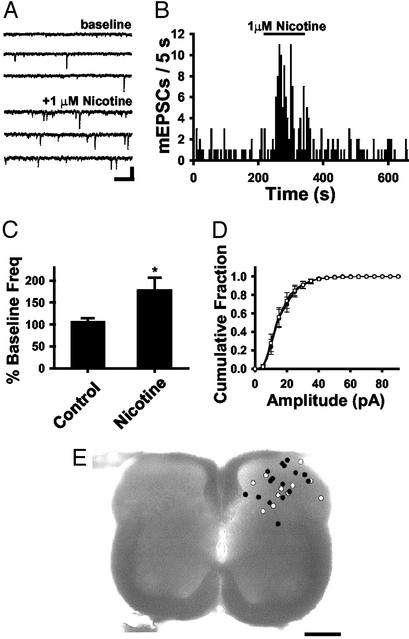
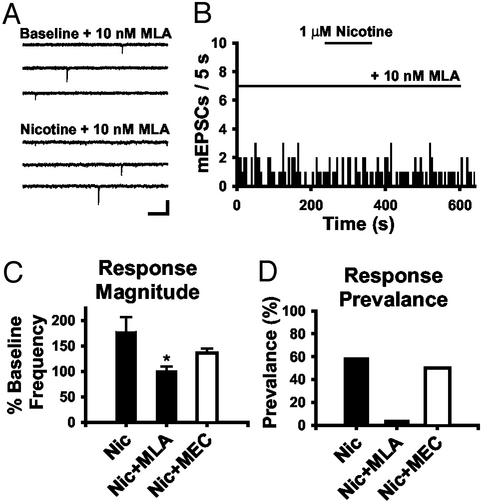
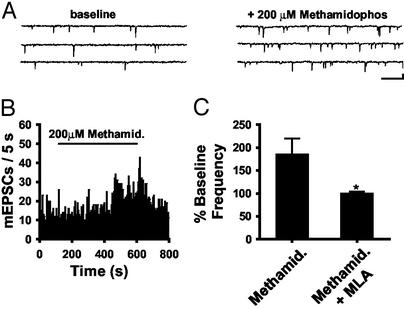
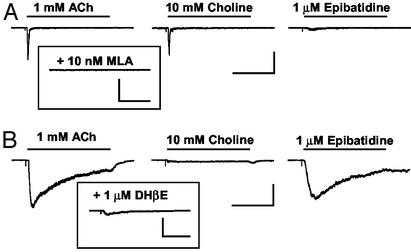
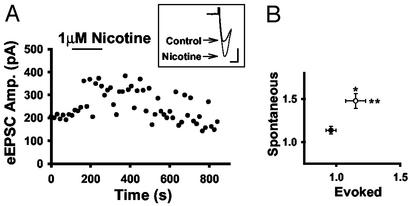
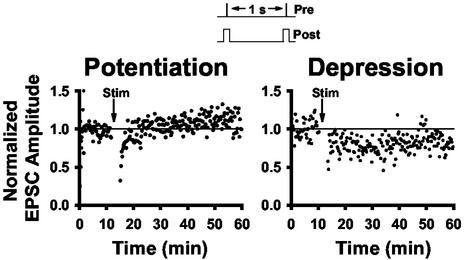
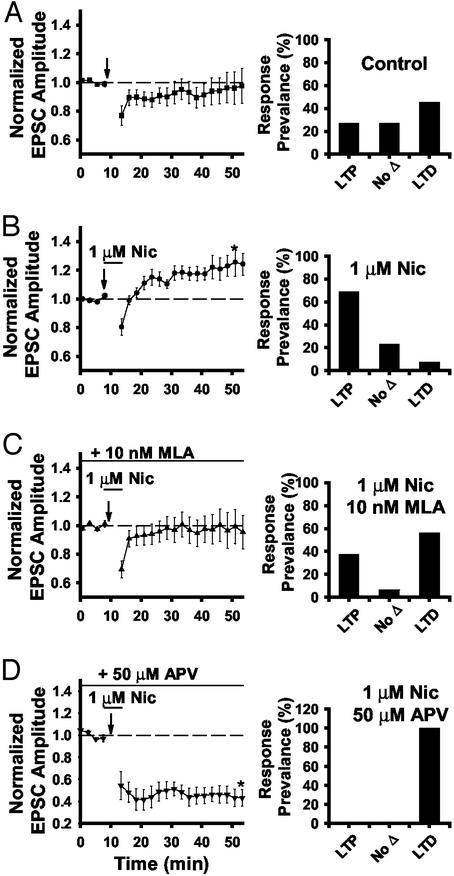
Spinal administration of nicotinic agonists can produce both hyperalgesic and analgesic effects in vivo. The cellular mechanisms underlying these behavioral phenomena are not understood. As a possible explanation for nicotinic hyperalgesia, we tested whether nicotinic acetylcholine receptors (nAChRs) could enhance excitatory transmission onto spinal cord dorsal horn neurons. Whole-cell patch-clamp recordings were performed in neonatal rat spinal cord slices. Activation of nAChRs enhanced glutamatergic synaptic transmission in 59% of dorsal horn neurons tested, and this effect was blocked by methyllycaconitine (10 nM), suggesting a key role for alpha7 nAChRs. Inhibition of acetylcholinesterase with methamidophos also enhanced transmission, demonstrating a similar effect of endogenous acetylcholine. nAChR activation also enhanced transmission by dorsal root entry zone stimulation, suggesting that alpha7 nAChRs on the central terminals of DRG afferents mediate this effect. Paired pre- and postsynaptic stimulation induced long-term potentiation of excitatory inputs to some of the dorsal horn neurons. Long-term potentiation induction was much more prevalent when nicotine was applied during stimulation. This effect also depended on both alpha7 nAChRs and N-methyl-d-aspartate glutamate receptors. Our findings demonstrate that alpha7 nAChRs can contribute to both short- and long-term enhancement of glutamatergic synaptic transmission in the spinal cord dorsal horn and provide a possible mechanism for nicotinic hyperalgesia.
Figures
Similar articles
-
Nicotinic modulation of GABAergic synaptic transmission in the spinal cord dorsal horn.Brain Res. 2005 Jan 21;1031(2):229-37. doi: 10.1016/j.brainres.2004.10.042. Brain Res. 2005. PMID: 15649448
-
Synaptic modulation of excitatory synaptic transmission by nicotinic acetylcholine receptors in spinal ventral horn neurons.Neuroscience. 2015 Apr 2;290:18-30. doi: 10.1016/j.neuroscience.2015.01.015. Epub 2015 Jan 19. Neuroscience. 2015. PMID: 25613686
-
The activation of nicotinic acetylcholine receptors enhances the inhibitory synaptic transmission in the deep dorsal horn neurons of the adult rat spinal cord.Mol Pain. 2007 Sep 25;3:26. doi: 10.1186/1744-8069-3-26. Mol Pain. 2007. PMID: 17894865 Free PMC article.
-
Repetitive stimulation induced potentiation of excitatory transmission in the rat dorsal horn: an in vitro study.J Neurophysiol. 1994 Jan;71(1):216-28. doi: 10.1152/jn.1994.71.1.216. J Neurophysiol. 1994. PMID: 7908954
-
Tonic inhibitory role of alpha4beta2 subtype of nicotinic acetylcholine receptors on nociceptive transmission in the spinal cord in mice.Pain. 2006 Nov;125(1-2):125-35. doi: 10.1016/j.pain.2006.05.011. Epub 2006 Jun 15. Pain. 2006. PMID: 16781069
Cited by
-
Ca2+ permeability of nicotinic acetylcholine receptors from rat dorsal root ganglion neurones.J Physiol. 2005 May 15;565(Pt 1):219-28. doi: 10.1113/jphysiol.2005.084871. Epub 2005 Mar 10. J Physiol. 2005. PMID: 15760934 Free PMC article.
-
Nicotinic acetylcholine receptors in neuropathic and inflammatory pain.FEBS Lett. 2018 Apr;592(7):1045-1062. doi: 10.1002/1873-3468.12884. Epub 2017 Oct 27. FEBS Lett. 2018. PMID: 29030971 Free PMC article. Review.
-
Inhibition of the regulator of G protein signalling RGS4 in the spinal cord decreases neuropathic hyperalgesia and restores cannabinoid CB1 receptor signalling.Br J Pharmacol. 2015 Nov;172(22):5333-46. doi: 10.1111/bph.13324. Epub 2015 Oct 25. Br J Pharmacol. 2015. PMID: 26478461 Free PMC article.
-
Molecular, Cellular and Circuit Basis of Cholinergic Modulation of Pain.Neuroscience. 2018 Sep 1;387:135-148. doi: 10.1016/j.neuroscience.2017.08.049. Epub 2017 Sep 8. Neuroscience. 2018. PMID: 28890048 Free PMC article. Review.
-
Nicotinic modulation of descending pain control circuitry.Pain. 2017 Oct;158(10):1938-1950. doi: 10.1097/j.pain.0000000000000993. Pain. 2017. PMID: 28817416 Free PMC article.
References
-
- Spande, T. F., Garraffo, H. M., Edwards, M. W., Yeh, H. J. C., Pannell, L. & Daly, J. W. (1992) J. Am. Chem. Soc. 114, 3475-3478.
-
- Bannon, A. W., Decker, M. W., Holladay, M. W., Curzon, P., Donnelly-Roberts, D., Puttfarcken, P. S., Bitner, R. S., Diaz, A., Dickenson, A. H., Porsolt, R. D., et al. (1998) Science 279, 77-81. - PubMed
-
- Flores, C. M. (2000) Pain 88, 1-6. - PubMed
-
- Decker, M. W. & Meyer, M. D. (1999) Biochem. Pharmacol. 58, 917-923. - PubMed
-
- Marubio, L. M., del Mar Arroyo-Jimenez, M., Cordero-Erausquin, M., Lena, C., Le Novere, N., de Kerchove d'Exaerde, A., Huchet, M., Damaj, M. I. & Changeux, J. P. (1999) Nature 398, 805-810. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
