

A mechanistic role for cardiac myocyte apoptosis in heart failure
- PMID: 12750399
- PMCID: PMC155051
- DOI: 10.1172/JCI17664
A mechanistic role for cardiac myocyte apoptosis in heart failure
Abstract
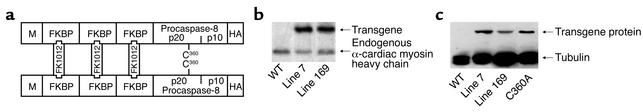
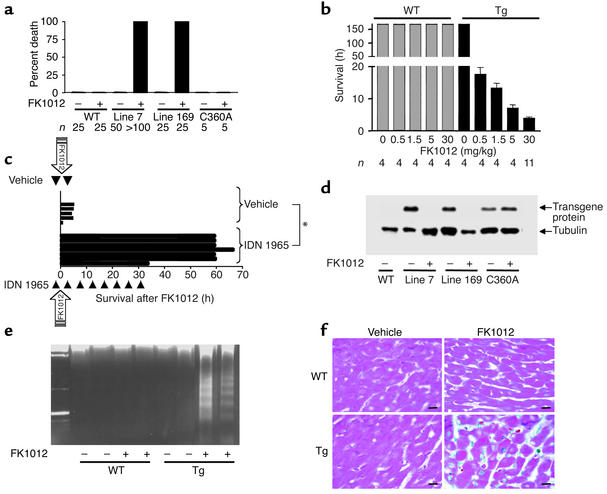
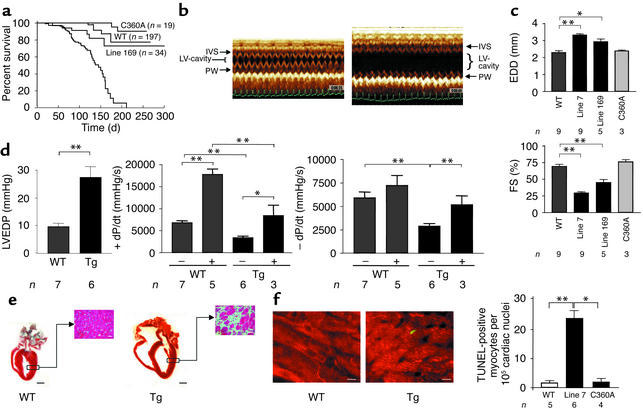
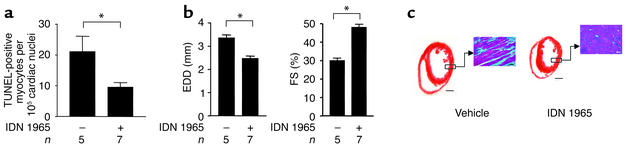
Heart failure is a common, lethal condition whose pathogenesis is poorly understood. Recent studies have identified low levels of myocyte apoptosis (80-250 myocytes per 10(5) nuclei) in failing human hearts. It remains unclear, however, whether this cell death is a coincidental finding, a protective process, or a causal component in pathogenesis. Using transgenic mice that express a conditionally active caspase exclusively in the myocardium, we demonstrate that very low levels of myocyte apoptosis (23 myocytes per 10(5) nuclei, compared with 1.5 myocytes per 10(5) nuclei in controls) are sufficient to cause a lethal, dilated cardiomyopathy. Interestingly, these levels are four- to tenfold lower than those observed in failing human hearts. Conversely, inhibition of cardiac myocyte death in this murine model largely prevents the development of cardiac dilation and contractile dysfunction, the hallmarks of heart failure. To our knowledge, these data provide the first direct evidence that myocyte apoptosis may be a causal mechanism of heart failure, and they suggest that inhibition of this cell death process may constitute the basis for novel therapies.
Figures
Comment in
-
A matter of life and death: cardiac myocyte apoptosis and regeneration.J Clin Invest. 2003 May;111(10):1457-9. doi: 10.1172/JCI18611. J Clin Invest. 2003. PMID: 12750394 Free PMC article. No abstract available.
References
-
- Braunwald E, Bristow MR. Congestive heart failure: fifty years of progress. Circulation. 2000;102:IV14–IV23. - PubMed
-
- Chien KR. Stress pathways and heart failure. Cell. 1999;98:555–558. - PubMed
-
- Lefkowitz RJ, Rockman HA, Koch WJ. Catecholamines, cardiac beta-adrenergic receptors, and heart failure. Circulation. 2000;101:1634–1637. - PubMed
-
- Marks AR. Ryanodine receptors, FKBP12, and heart failure. Front. Biosci. 2002;7:d970–d977. - PubMed
-
- Luo W, et al. Targeted ablation of the phospholamban gene is associated with markedly enhanced myocardial contractility and loss of beta-agonist stimulation. Circ. Res. 1994;75:401–409. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
