

doi: 10.1128/JB.185.11.3453-3457.2003.
Conserved eukaryotic histone-fold residues substituted into an archaeal histone increase DNA affinity but reduce complex flexibility
Affiliations
- PMID: 12754245
- PMCID: PMC155370
- DOI: 10.1128/JB.185.11.3453-3457.2003
Item in Clipboard
Conserved eukaryotic histone-fold residues substituted into an archaeal histone increase DNA affinity but reduce complex flexibility
J Bacteriol.
2003 Jun.
Abstract
Although the archaeal and eukaryotic nucleosome core histones evolved from a common ancestor, conserved lysine residues are present at DNA-binding locations in all four eukaryotic histones that are not present in the archaeal histones. Introduction of lysine residues at the corresponding locations into an archaeal histone, HMfB, generated a variant with increased affinity for DNA that formed more compact complexes with DNA. However, these complexes no longer facilitated the circularization of short DNA molecules and had lost the flexibility to wrap DNA alternatively in either a negative or positive supercoil.
Figures
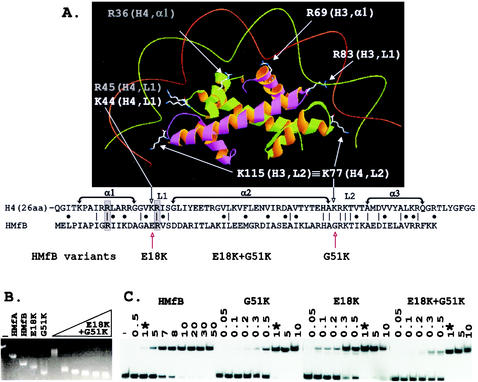
Histone fold-DNA complex, sequence alignment, and electrophoretic mobility shift assays of complexes formed by HMfB variants. (A) The histone folds of a histone H3-H4 heterodimer with conserved side chains positioned to bind DNA as in the eukaryotic nucleosome (7, 8). The side chains of lysines conserved at the same location in L2 in all four eukaryotic histones, illustrated in the figure by K115 in H3 and K77 in H4, extend the contacts of each histone fold dimer by one-half helical turn in each direction (8). As identified in the sequence alignment, the corresponding histone fold position is occupied by G51 in HMfB and by a glycine in almost all archaeal histones (16). The sequences that form α1, α2, α3, L1, and L2 of the histone folds of H4 and HMfB are indicated, with identical residues and conservative differences identified by vertical lines and dots, respectively. Conserved arginines located in α1 and L1 that also extend side chains to make DNA contacts (7, 8) are boxed and shaded. (B) Agarose gel electrophoresis of linear pBR322 (lane −) and of the complexes formed at 25°C by wild-type HMfA and HMfB (15, 16) and the HMfB, E18K, and G51K variants (18) at saturating histone-to-pBR322 DNA ratios is shown; adjacent lanes show the complexes formed by assembly of the HMfB E18K+G51K variant at histone-to-pBR322 DNA mass ratios of 1, 3, 4, 5, 6, and 7. The histone-DNA assembly conditions, agarose gel electrophoresis, and ethidium bromide staining protocols have been described in detail previously (15). (C) Autoradiogram of the polyacrylamide gel electrophoretic separations of the complexes formed at 25°C in reaction mixtures that contained 0.1 ng of 32P-labeled 110-bp clone 20 DNA (3), 1 ng of competitor sonicated herring sperm DNA, and the amounts (in nanograms) of HMfB or the HMfB variant listed above the gel. Lane −, protein-free DNA control; ∗, the lanes that contained the complexes formed under identical conditions by 1 ng of HMfB or 1 ng of a HMfB variant. The histone-DNA assembly conditions, polyacrylamide gel electrophoresis, and autoradiography procedures used have been described previously (3, 4).
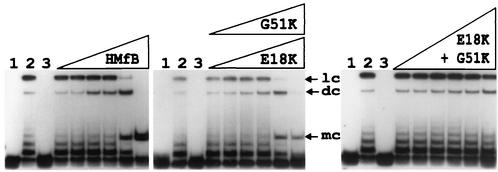
Autoradiogram of the electrophoretic separation of the DNA products generated by incubation of HMfB and the HMfB variants with an 88-bp DNA and DNA ligase. The 88-bp DNA molecule was generated by SpeI and XbaI digestion of pLITMUS28Δ10; it was 32P labeled and used in circularization assays as previously described (2, 15). Lane 1, 0.5-ng aliquot of the 32P-labeled DNA; lane 2, products of incubation of this DNA with T4 DNA ligase for 12 h at 16°C in the absence of histones; lane 3, products of incubation of an aliquot of this DNA with 10 ng of HMfB or an HMfB variant followed by deproteinization without exposure to DNA ligase. The adjacent lanes contained the DNA products resulting from reaction mixtures that contained 0.5 ng of DNA and 0.5, 1, 3, 5, 10, or 25 ng of the indicated histone; the products were incubated for 12 h at 16°C with T4 DNA ligase and then subjected to deproteinization. Almost identical results were obtained with the E18K (shown) and G51K variants. Linear concatemer (lc), dimer circle (dc), and monomer circle (mc) ligation products are identified (3). Note that monomer circles were not generated when aliquots of the DNA were incubated with DNA ligase in the absence of an archaeal histone (lanes 2).
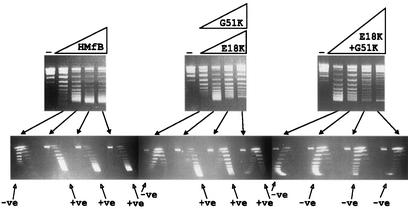
One- and two-dimensional agarose gel electrophoretic separations of the topoisomers generated by HMfB or HMfB variants assembled on relaxed, circular pUC18 DNA molecules. The procedures used to assemble complexes, remove plectonemic supercoils, deproteinize, and separate pUC18 topoisomers by one- and two-dimensional agarose gel electrophoresis have been described previously (9, 10). The control lanes (−) contained relaxed pUC18 DNA; adjacent lanes contained the pUC18 topoisomers generated by HMfB or HMfB variant assembly on aliquots of this DNA at histone-to-DNA mass ratios of 0.4, 0.6, 0.8, and 1. Almost identical results were obtained with the E18K variant (shown) and G51 variant. Topoisomers were separated in the first dimension (upper gels) on the basis of linking number, with increasing linking number resulting in increasing mobility, and in the second dimension (lower gels) on the basis of negative (−ve) or positive (+ve) supercoiling. Nicked and relaxed circular DNAs migrated together and formed the band near the top of each gel.
Similar articles
-
Mutational analysis of archaeal histone-DNA interactions.J Mol Biol. 2000 Mar 17;297(1):39-47. doi: 10.1006/jmbi.2000.3546. J Mol Biol. 2000. PMID: 10704305
-
Archaeal histones: structures, stability and DNA binding.Biochem Soc Trans. 2004 Apr;32(Pt 2):227-30. doi: 10.1042/bst0320227. Biochem Soc Trans. 2004. PMID: 15046577 Review.
-
Archaeal histones and the origin of the histone fold.Curr Opin Microbiol. 2006 Oct;9(5):520-5. doi: 10.1016/j.mib.2006.08.003. Epub 2006 Aug 22. Curr Opin Microbiol. 2006. PMID: 16920388
-
Archaeal histone tetramerization determines DNA affinity and the direction of DNA supercoiling.J Biol Chem. 2002 Aug 23;277(34):30879-86. doi: 10.1074/jbc.M203674200. Epub 2002 Jun 10. J Biol Chem. 2002. PMID: 12058041
-
Histones and nucleosomes in Archaea and Eukarya: a comparative analysis.Extremophiles. 1998 Aug;2(3):141-8. doi: 10.1007/s007920050053. Extremophiles. 1998. PMID: 9783158 Review.
Cited by
-
Old cogs, new tricks: the evolution of gene expression in a chromatin context.Nat Rev Genet. 2019 May;20(5):283-297. doi: 10.1038/s41576-019-0105-7. Nat Rev Genet. 2019. PMID: 30886348 Review.
-
Structure and function of archaeal histones.PLoS Genet. 2018 Sep 13;14(9):e1007582. doi: 10.1371/journal.pgen.1007582. eCollection 2018 Sep. PLoS Genet. 2018. PMID: 30212449 Free PMC article. Review.
-
Histone variants in archaea and the evolution of combinatorial chromatin complexity.Proc Natl Acad Sci U S A. 2020 Dec 29;117(52):33384-33395. doi: 10.1073/pnas.2007056117. Epub 2020 Dec 7. Proc Natl Acad Sci U S A. 2020. PMID: 33288720 Free PMC article.
-
The hydrophobicity of the H3 histone fold differs from the hydrophobicity of the other three folds.J Mol Evol. 2005 Mar;60(3):354-64. doi: 10.1007/s00239-004-0193-6. J Mol Evol. 2005. PMID: 15871046
References
-
- Bailey, K. A., S. L. Pereira., J. Widom, and J. N. Reeve. 2000. Archaeal histone selection of nucleosome positioning sequences and the procaryotic origin of histone-dependent genome evolution. J. Mol. Biol. 303:25-34. - PubMed
-
- Bailey, K. A., F. Marc, K. Sandman, and J. N. Reeve. 2002. Both DNA and histone fold sequences contribute to archaeal nucleosome stability. J. Biol. Chem. 277:9293-9301. - PubMed
-
- Bell, S. D., C. H. Botting, B. N. Wardleworth, S. P. Jackson, and M. F. White. 2002. The interaction of Alba, a conserved archaeal chromatin protein, with Sir2 and its regulation by acetylation. Science 296:148-151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
