

An uncapped RNA suggests a model for Caenorhabditis elegans polycistronic pre-mRNA processing
- PMID: 12756326
- PMCID: PMC1370435
- DOI: 10.1261/rna.2128903
An uncapped RNA suggests a model for Caenorhabditis elegans polycistronic pre-mRNA processing
Abstract
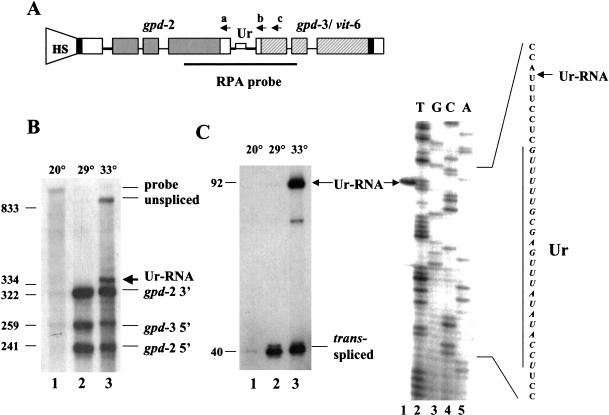
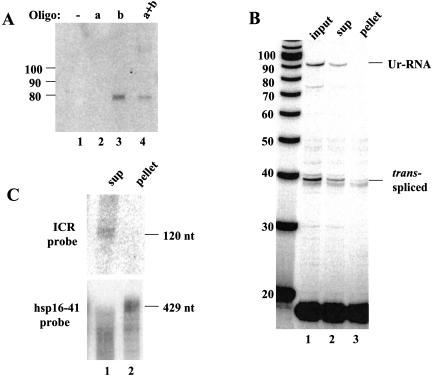
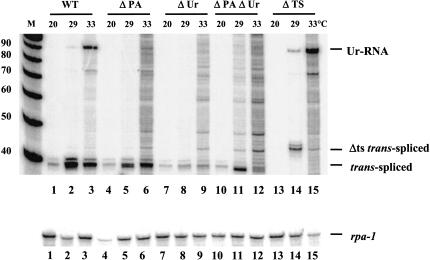
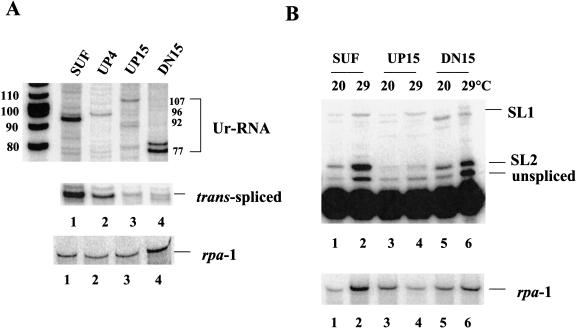
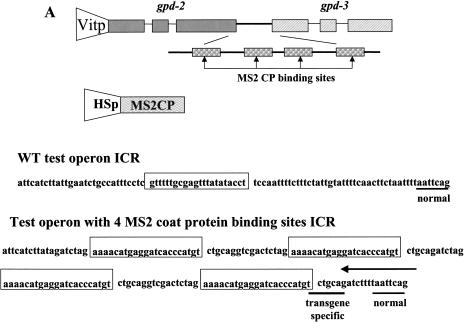
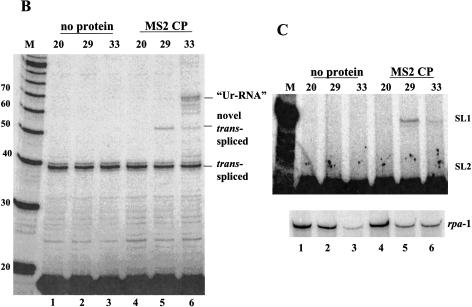
Polycistronic pre-mRNAs from Caenohabditis elegans operons are processed by internal cleavage and polyadenylation to create 3' ends of mature mRNAs. This is accompanied by trans-splicing with SL2 approximately 100 nucleotides downstream of the 3' end formation sites to create the 5' ends of downstream mRNAs. SL2 trans-splicing depends on a U-rich element (Ur), located approximately 70 nucleotides upstream of the trans-splice site in the intercistronic region (ICR), as well as a functional 3' end formation signal. Here we report the existence of a novel gene-length RNA, the Ur-RNA, starting just upstream of the Ur element. The expression of Ur-RNA is dependent on 3' end formation as well as on the presence of the Ur element, but does not require a trans-splice site. The Ur-RNA is not capped, and alteration of the location of the Ur element in either the 5' or 3' direction alters the location of the 5' end of the Ur-RNA. We propose that a 5' to 3' exonuclease degrades the precursor RNA following cleavage at the poly(A) site, stopping when it reaches the Ur element, presumably attributable to a bound protein. Part of the function of this protein can be performed by the MS2 coat protein. Recruitment of coat protein to the ICR in the absence of the Ur element results in accumulation of an RNA equivalent to Ur-RNA, and restores trans-splicing. Only SL1, however, is used. Therefore, coat protein is sufficient for blocking the exonuclease and thereby allowing formation of a substrate for trans-splicing, but it lacks the ability to recruit the SL2 snRNP. Our results also demonstrate that MS2 coat protein can be used as an in vivo block to an exonuclease, which should have utility in mRNA stability studies.
Figures
Similar articles
-
Trans-splicing and operons in C. elegans.WormBook. 2012 Nov 20:1-11. doi: 10.1895/wormbook.1.5.2. WormBook. 2012. PMID: 23175478 Free PMC article. Review.
-
C. elegans sequences that control trans-splicing and operon pre-mRNA processing.RNA. 2007 Sep;13(9):1409-26. doi: 10.1261/rna.596707. Epub 2007 Jul 13. RNA. 2007. PMID: 17630324 Free PMC article.
-
Polycistronic pre-mRNA processing in vitro: snRNP and pre-mRNA role reversal in trans-splicing.Genes Dev. 2010 Aug 1;24(15):1645-58. doi: 10.1101/gad.1940010. Epub 2010 Jul 12. Genes Dev. 2010. PMID: 20624853 Free PMC article.
-
Intercistronic region required for polycistronic pre-mRNA processing in Caenorhabditis elegans.Mol Cell Biol. 2001 Feb;21(4):1111-20. doi: 10.1128/MCB.21.4.1111-1120.2001. Mol Cell Biol. 2001. PMID: 11158298 Free PMC article.
-
Trans-splicing and operons.WormBook. 2005 Jun 25:1-9. doi: 10.1895/wormbook.1.5.1. WormBook. 2005. PMID: 18050426 Review.
Cited by
-
Upf1 ATPase-dependent mRNP disassembly is required for completion of nonsense- mediated mRNA decay.Cell. 2010 Dec 10;143(6):938-50. doi: 10.1016/j.cell.2010.11.043. Cell. 2010. PMID: 21145460 Free PMC article.
-
A novel family of C. elegans snRNPs contains proteins associated with trans-splicing.RNA. 2007 Apr;13(4):511-20. doi: 10.1261/rna.426707. Epub 2007 Feb 5. RNA. 2007. PMID: 17283210 Free PMC article.
-
RNA polymerase II CTD S2P is dispensable for embryogenesis but mediates exit from developmental diapause in C. elegans.Sci Adv. 2020 Dec 9;6(50):eabc1450. doi: 10.1126/sciadv.abc1450. Print 2020 Dec. Sci Adv. 2020. PMID: 33298437 Free PMC article.
-
High frequency trans-splicing in a cell line producing spliced and polyadenylated RNA polymerase I transcripts from an rDNA-myc chimeric gene.Nucleic Acids Res. 2005 Apr 22;33(7):2332-42. doi: 10.1093/nar/gki530. Print 2005. Nucleic Acids Res. 2005. PMID: 15849319 Free PMC article.
-
Trans-splicing and operons in C. elegans.WormBook. 2012 Nov 20:1-11. doi: 10.1895/wormbook.1.5.2. WormBook. 2012. PMID: 23175478 Free PMC article. Review.
References
-
- Blumenthal, T., Evans, D., Link, C.D., Guffanti, A., Lawson, D., Thierry-Mieg, J., Thierry-Mieg, D., Chiu, W.L., Duke, K., Kiraly, M., et al. 2002. A global analysis of Caenorhabditis elegans operons. Nature 417: 851–854. - PubMed
-
- Blumenthal, T. and Spieth, J. 1996. Gene structure and organization in Caenorhabditis elegans. Curr. Opin. Genet. Dev. 6: 692–698. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources