

Review
doi: 10.1046/j.1365-2567.2003.01657.x.
Innate self recognition by an invariant, rearranged T-cell receptor and its immune consequences
Affiliations
- PMID: 12757612
- PMCID: PMC1782955
- DOI: 10.1046/j.1365-2567.2003.01657.x
Item in Clipboard
Review
Innate self recognition by an invariant, rearranged T-cell receptor and its immune consequences
Immunology.
2003 Jun.
Abstract
This review attempts to illuminate the glycolipid antigen presentation properties of CD1d, how CD1d controls the function of natural T (iNKT) cells and how CD1d and iNKT cells interact to jump-start the immune system. It is postulated that the CD1d-iNKT cell system functions as a sensor, sensing alterations in cellular lipid content by virtue of its affinity for such ligands. The presentation of a neo-self glycolipid, presumably by infectious assault of antigen-presenting cells, activates iNKT cells, which promptly release pro-inflammatory and anti-inflammatory cytokines and jump-start the immune system.
Figures
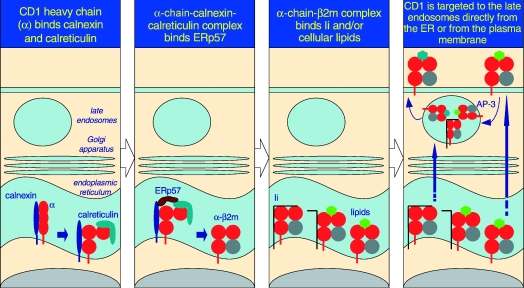
The predicted pathway for CD1d assembly and intracellular trafficking. As it is a type I integral membrane glycoprotein, the folding and assembly of CD1d occur in the rough ER. Here, calnexin, calreticulin and ERp57 assist the folding of the CD1d α-chain. β2m associates with the folded α-chain, unlike MHC class I molecules, which bind calreticulin only after association with the light chain. The α-chain+β2m+ complex forms a structure receptive to Ii and/or resident lipids and glycolipids. Upon stable association, the CD1d (α-chain and β2m)–glycolipid complexes egress from the ER and negotiate the secretory pathway to the plasma membrane. Because of the Yxxφ internalization sequence within the cytosolic tail of CD1d, it is rapidly internalized into the late endosomes in an AP-3-dependent manner and recycled back to the plasma membrane. In contrast, Ii-associated CD1d may be directly targeted to the late endosomes, and thence egress to the cell surface. During its time in the late endosomes, lipids bound to CD1d in the ER are exchanged for another, presumably cellular, glycolipid that is presented to iNKT cells.
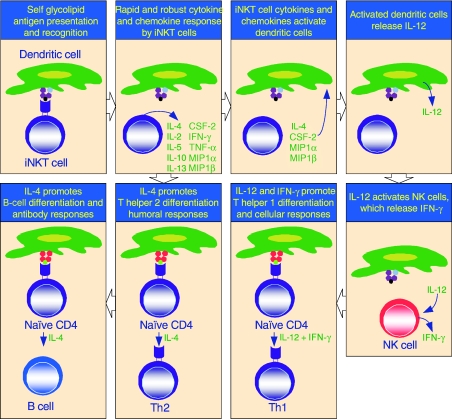
The immunological functions of iNKT cells. iNKT cells participate in cross talk between members of the innate and the adaptive immune system by deploying cytokine/chemokine messengers. Upon activation in vivo, iNKT cells rapidly secrete several cytokines/chemokines. Of these cytokines/chemokines, IL-4, CSF-2, MIP1α and MIP1β facilitate the recruitment, activation and differentiation of macrophages and dendritic cells resulting in the production of IL-12 and possibly other factors. IL-12, in turn, stimulates NK cells to secrete IFN-γ. Along with IL-12, IFN-γ can polarise the differentiation of antigen-activated CD4+ T cells towards a Th1 phenotype. IL-4 can skew the differentiation of CD4+ T cells towards a Th2 phenotype. IL-4 can also activate B cells in vivo resulting in their differentiation to antibody secreting-plasma cells. Thus iNKT cells have the potential for jump-starting an immune response.
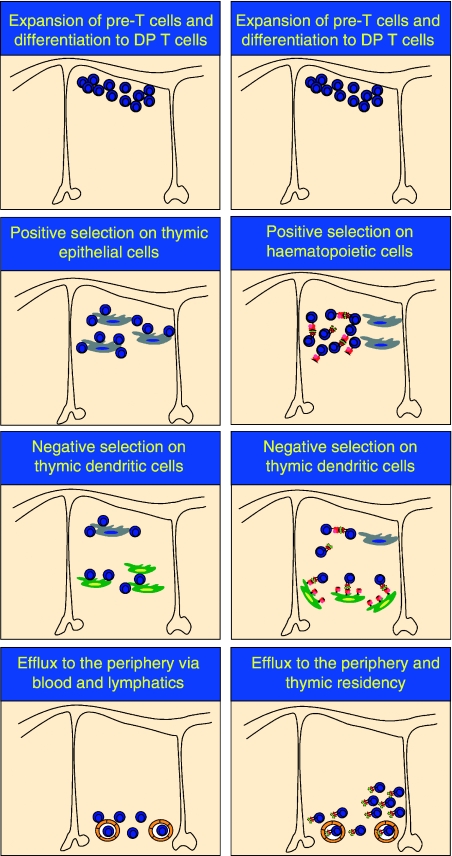
A schematic of cell-to-cell interactions during positive and negative selection in the thymus. Conventional T and iNKT cells develop in the thymus, and share DN T-cell precursors. However, positive selection of iNKT cells proceeds via interactions with haematopietic cells, while conventional T cells require thymic epithelium for the positive developmental signal. Negative selection of both conventional T and iNKT cells is mediated by thymic dendritic cells. Both conventional T and iNKT cells are exported to the periphery following successful thymic education; however, a subset of iNKT cells remain thymic residents. Thus, conventional T and iNKT cells have distinct cellular requirements during positive and negative selection.
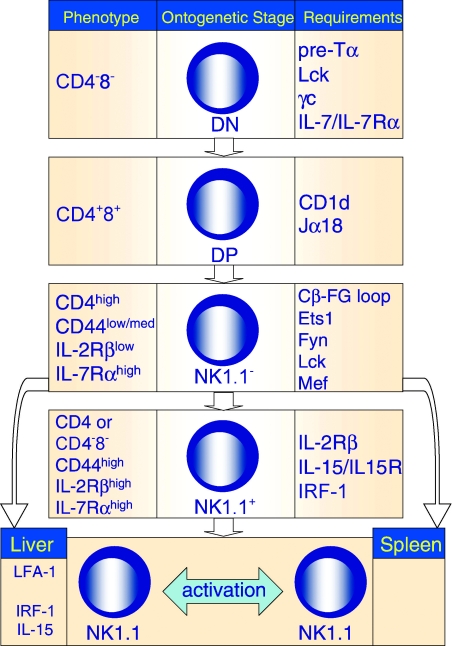
Molecular control of iNKT cell development. The development of a functional repertoire of iNKT cells requires progression through several developmental intermediates in the thymus, export to the periphery and homeostatic proliferation in resident organs. In the column on the right are molecular factors required for iNKT cell progression past the stage shown in the middle column. The phenotype of the developmental intermediate is shown in the left column.
Similar articles
-
CD1d and natural T cells: how their properties jump-start the immune system.Cell Mol Life Sci. 2001 Mar;58(3):442-69. doi: 10.1007/PL00000869. Cell Mol Life Sci. 2001. PMID: 11315191 Free PMC article. Review.
-
CD1d-restricted glycolipid antigens: presentation principles, recognition logic and functional consequences.Expert Rev Mol Med. 2008 Jul 7;10:e20. doi: 10.1017/S1462399408000732. Expert Rev Mol Med. 2008. PMID: 18601810 Review.
-
Therapeutic potential of CD1d-restricted invariant natural killer T cell-based treatment for autoimmune diseases.Int Rev Immunol. 2007 Jan-Apr;26(1-2):73-94. doi: 10.1080/08830180601070252. Int Rev Immunol. 2007. PMID: 17454265 Review.
-
Characterization and functional analysis of mouse invariant natural T (iNKT) cells.Curr Protoc Immunol. 2006 Jul;Chapter 14:14.13.1-14.13.27. doi: 10.1002/0471142735.im1413s73. Curr Protoc Immunol. 2006. PMID: 18432968
-
Cutting edge: nonglycosidic CD1d lipid ligands activate human and murine invariant NKT cells.J Immunol. 2008 May 15;180(10):6452-6. doi: 10.4049/jimmunol.180.10.6452. J Immunol. 2008. PMID: 18453560
Cited by
-
Beta-glycoglycosphingolipid-induced alterations of the STAT signaling pathways are dependent on CD1d and the lipid raft protein flotillin-2.Am J Pathol. 2009 Apr;174(4):1390-9. doi: 10.2353/ajpath.2009.080841. Epub 2009 Feb 26. Am J Pathol. 2009. PMID: 19246642 Free PMC article.
-
Commitment toward the natural T (iNKT) cell lineage occurs at the CD4+8+ stage of thymic ontogeny.Proc Natl Acad Sci U S A. 2005 Apr 5;102(14):5114-9. doi: 10.1073/pnas.0408449102. Epub 2005 Mar 25. Proc Natl Acad Sci U S A. 2005. PMID: 15792999 Free PMC article.
-
Alpha versus beta: are we on the way to resolve the mystery as to which is the endogenous ligand for natural killer T cells?Clin Exp Immunol. 2009 Dec;158(3):300-7. doi: 10.1111/j.1365-2249.2009.04030.x. Epub 2009 Sep 30. Clin Exp Immunol. 2009. PMID: 19793337 Free PMC article.
References
-
- Bendelac A, Bonneville M, Kearney JF. Autoreactivity by design: innate B and T lymphocytes. Nature Rev Immunol. 2001;1:177–86. - PubMed
-
- Smyth MJ, Crowe NY, Hayakawa Y, Takeda K, Yagita H, Godfrey DI. NKT cells – conductors of tumor immunity? Curr Opin Immunol. 2002;14:165–71. - PubMed
-
- Hong S, Wilson MT, Serizawa I, et al. The natural killer T-cell ligand α-galactosylceramide prevents autoimmune diabetes in non-obese diabetic mice. Nature Med. 2001;7:1052–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
