

Review
doi: 10.1523/JNEUROSCI.23-10-03972.2003.
Brain function and neurophysiological correlates of signals used in functional neuroimaging
Affiliations
- PMID: 12764081
- PMCID: PMC6741101
- DOI: 10.1523/JNEUROSCI.23-10-03972.2003
Item in Clipboard
Review
Brain function and neurophysiological correlates of signals used in functional neuroimaging
J Neurosci.
.
No abstract available
Figures
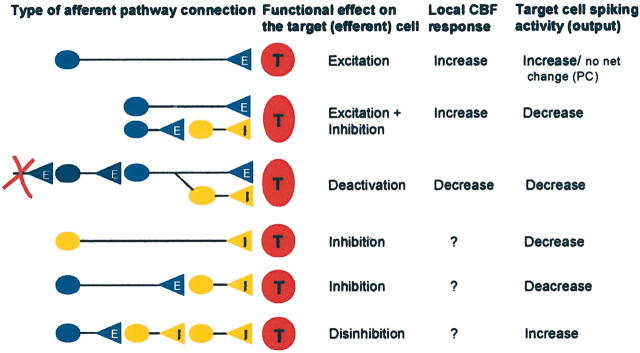
Schematic drawings of neuronal circuits thought to be relevant for studies of the coupling between neuronal activity and CBF or BOLD signals. Our studies and those of others have demonstrated the hemodynamic changes produced by monosynaptic excitation as shown in the first line, mixed disynaptic inhibition/excitation as shown in the second line, and deactivation as shown in the third line. The bottom three panels show other categories of neuronal circuits that will need to be examined to obtain a better understanding of the neuronal basis of CBF or BOLD signals. E, Excitatory; I, inhibitory; T, target cell.
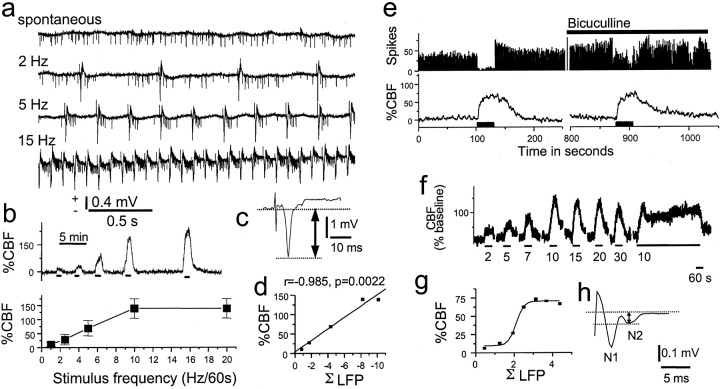
Interrelationship of action potentials, synaptic activity, and local changes in CBF in rat cerebellar cortex. a-d, Recordings of action potentials (a), CBF (b), LFP (c), and CBF versus ΣLFP (d) for stimulation of the climbing fiber system. e-h, Recordings of action potentials and CBF (e), CBF (f), ΣLFP versus CBF (g), and LFP (h). a, Under control conditions Purkinje cells discharges were mainly simple action potentials that were triggered by the mossy-fiber granule cell system. In response to climbing fiber stimulation, this spike pattern changed to become dominated by complex spikes at 1, 2, 5, 7, 10, and 15 Hz. However, the total number of spikes remained the same. b, CBF increased frequency dependency at the same time but already reached a plateau at 10 Hz, whereas the number of complex spikes continued to increase up to 20 Hz. This suggested that the rises in CBF and spike rate were dissociated. c, The LFP evoked by climbing fiber stimulation was monophasic and negative, produced by ion fluxes in postsynaptic cellular elements. d, CBF increases evoked by climbing fiber stimulation were linearly correlated to ΣLFP recorded at a depth of 400 μm, i.e., corresponding to maximal postsynaptic activity. e, Purkinje cell spike activity and CBF evoked by parallel fiber stimulation. Spikes almost disappeared at 1-3 sec after onset of stimulation, and the spontaneous firing did not return to normal until 19-25 sec after end of stimulation. CBF increased during stimulation, continued to increase for 5-10 sec after end of stimulation, and reversed to baseline after 40-50 sec. The GABAA antagonist bicuculline (0.5 mm , topical application; horizontal bar) did not affect CBF but attenuated the inhibition of Purkinje cell spike activity during stimulation. This figure demonstrates the complete dissociation of spikes and vascular signal. f, Frequency dependence of CBF rises in response to parallel fiber stimulation. g, Correlation of frequency-dependent increases in CBF to ΣLFP indicated a sigmoid relationship. h, Enlarged LFP to indicate how amplitudes were calculated. The first negativity (N1) was associated with the presynaptic action potential, whereas the second negativity (N2) represents postsynaptic excitation. The amplitude was measured as the voltage difference between the two dashed lines. Adapted from Mathiesen et al. (1998).
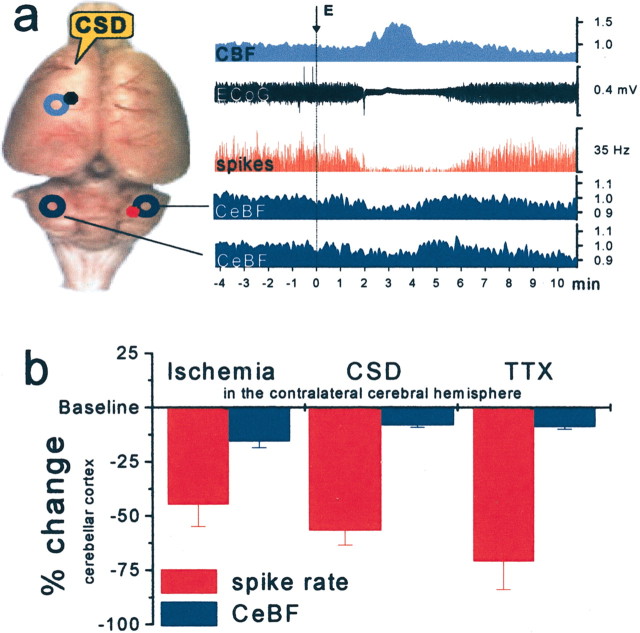
Deactivation produced by functional ablation of the left frontal cortex by an episode of cortical spreading depression (CSD) produced decreased activity in the cortico-pontine-cerebellar projections and in turn reduced spiking activity in cerebellar Purkinje cells in the contralateral cerebellar cortex. a, The rat brain viewed from above. CSD was elicited in the left frontal cortex (E, right) and identified by the suppression of electrocorticographic (ECoG) activity and the concomitant rise in CBF (top two recordings). At the same time, the spontaneous spiking (red, third recording) in Crus II of the contralateral cerebellar cortex became strongly attenuated, whereas CBF in the contralateral and ipsilateral cerebellar hemisphere decreased slightly (fourth and fifth recordings). y-axis without units indicates normalized data. b, Summary figure showing decreases in spontaneous Purkinje cell spike activity and baseline cerebellar CBF in response to impaired contralateral neocortical function attributable to focal ischemia (n = 10), CSD (n = 4), and TTX (n = 4). This figure illustrates the discrepancy between the large decreases in spontaneous neuronal activity and the small decreases in baseline cerebellar CBF during deactivation. Relative changes (%) of mean spike rate (red) and cerebellar blood flow (CeBF) (blue) are shown. Adapted from Gold and Lauritzen (2002).
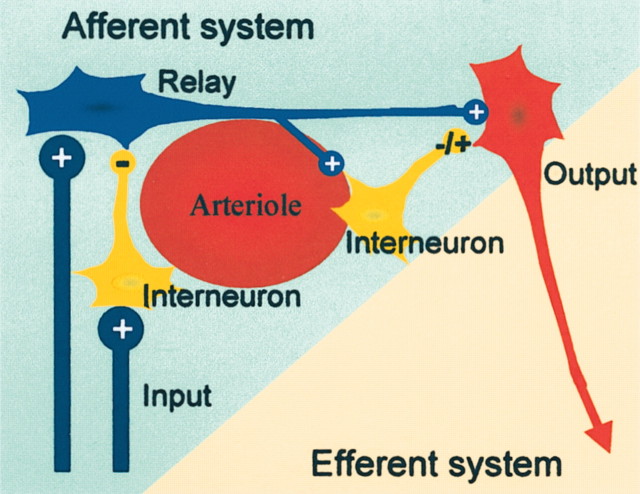
Neuronal network model thought to illustrate our working hypothesis of the neural basis of CBF and BOLD signals used in functional neuroimaging. Activity-dependent increases in BOLD or CBF are probably related to activity in the afferent input system, i.e., local excitatory and inhibitory synaptic activity in relay neurons, interneurons, and the efferent cell. Local spiking in horizontal projections of activated neurons may contribute as well, but this remains to be shown. Under most conditions increases in CBF are unrelated to spiking activity in efferent neuron. Therefore, it is not possible on the basis of a local increase in CBF (or BOLD) to decide whether the spiking activity out of an activated region is increased or decreased. Adapted from Lauritzen (2001).
References
-
- Akgoren N, Dalgaard P, Lauritzen M ( 1996) Cerebral blood flow increases evoked by electrical stimulation of rat cerebellar cortex: relation to excitatory synaptic activity and nitric oxide synthesis. Brain Res 710: 204–214. - PubMed
-
- Akgoren N, Mathiesen C, Rubin I, Lauritzen M ( 1997) Laminar analysis of activity-dependent increases of CBF in rat cerebellar cortex: dependence on synaptic strength. Am J Physiol 273: H1166–H1176. - PubMed
-
- Ances BM, Zarahn E, Greenberg JH, Detre JA ( 2000) Coupling of neural activation to blood flow in the somatosensory cortex of rats is time-intensity separable, but not linear. J Cereb Blood Flow Metab 20: 921–930. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical