

Exploitation of astrocytes by glioma cells to facilitate invasiveness: a mechanism involving matrix metalloproteinase-2 and the urokinase-type plasminogen activator-plasmin cascade
- PMID: 12764090
- PMCID: PMC6741112
- DOI: 10.1523/JNEUROSCI.23-10-04034.2003
Exploitation of astrocytes by glioma cells to facilitate invasiveness: a mechanism involving matrix metalloproteinase-2 and the urokinase-type plasminogen activator-plasmin cascade
Abstract
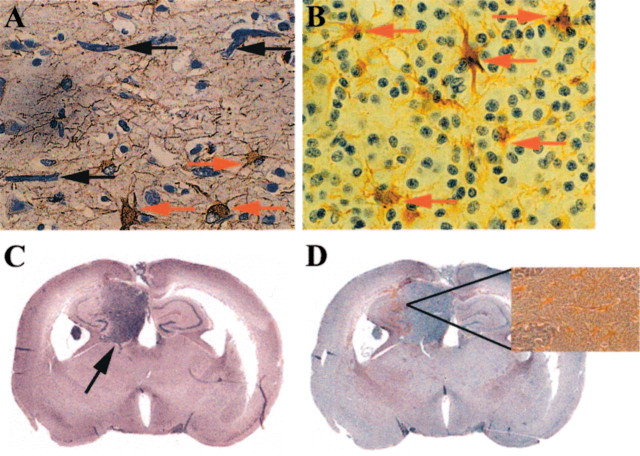
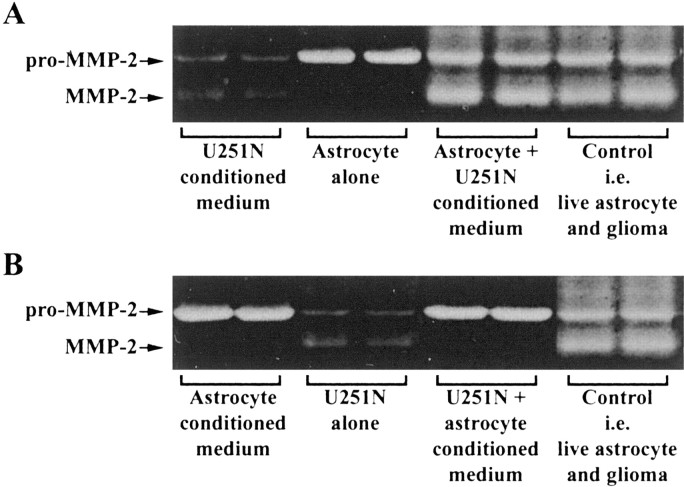
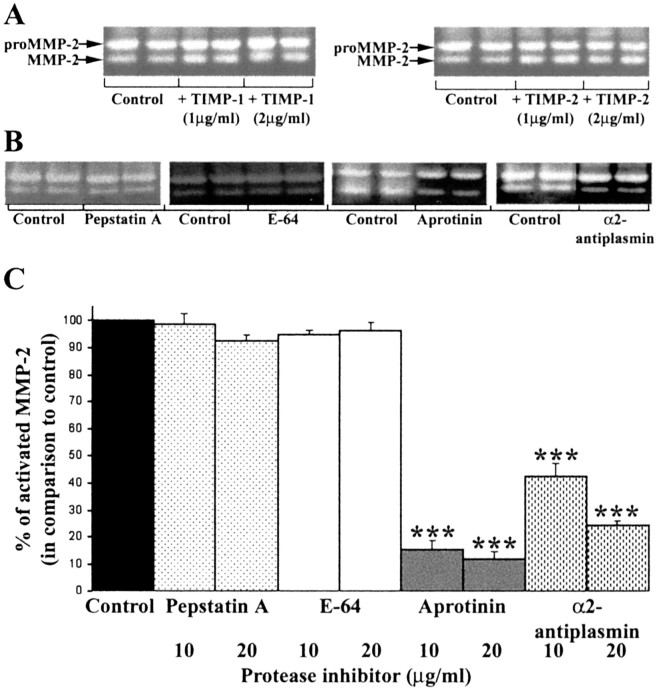
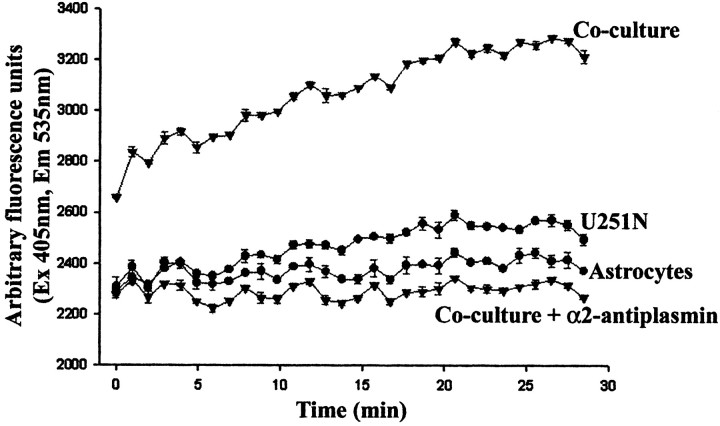
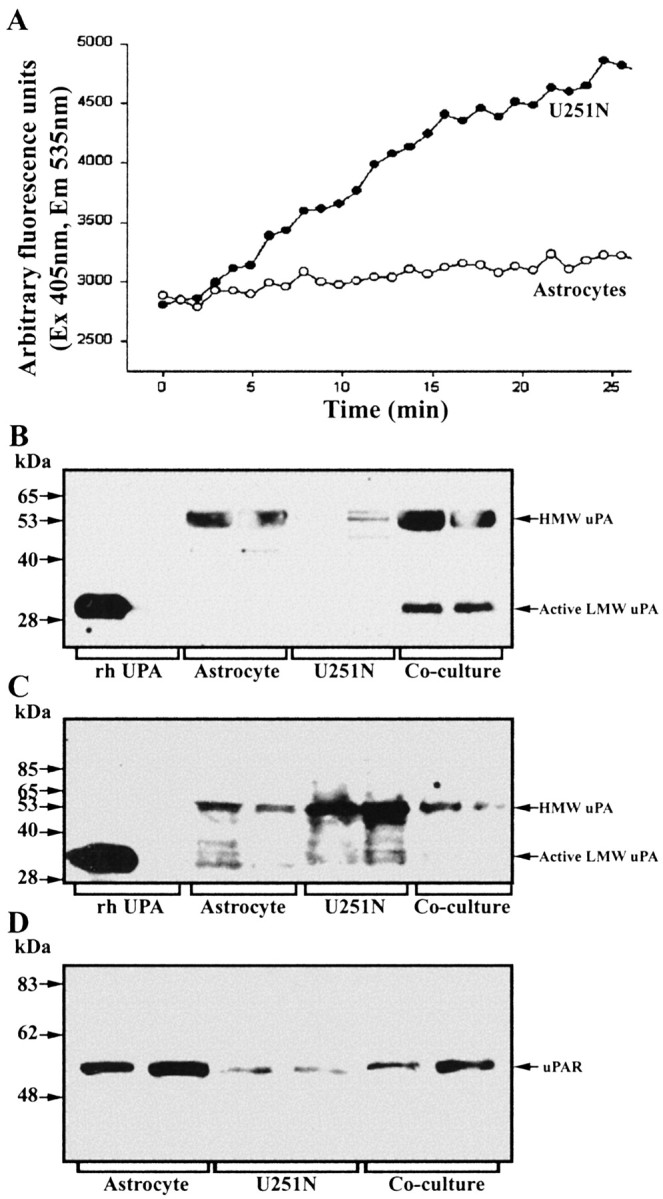
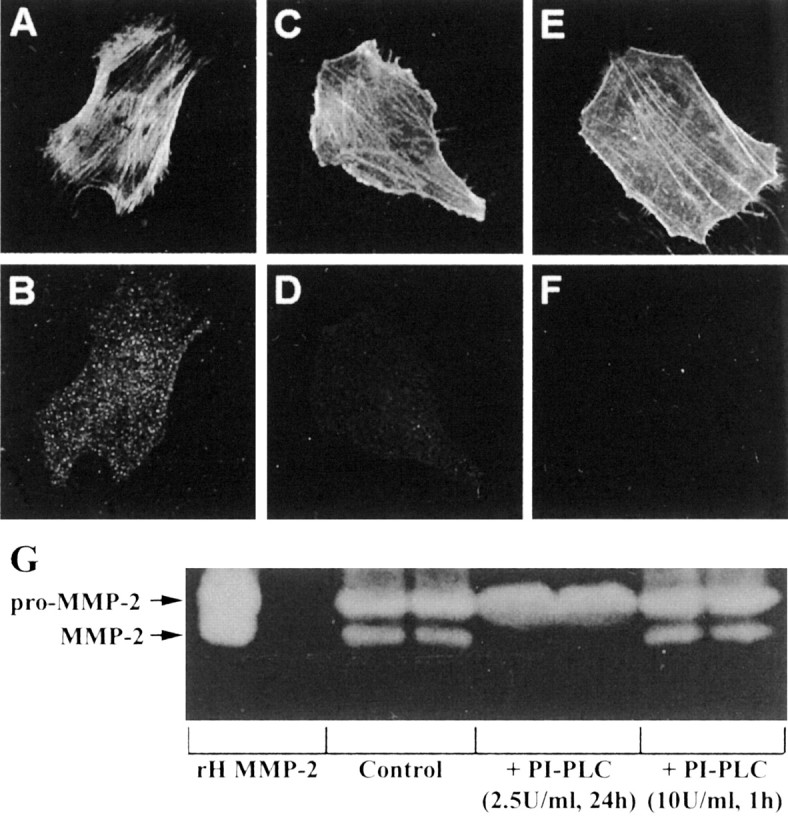
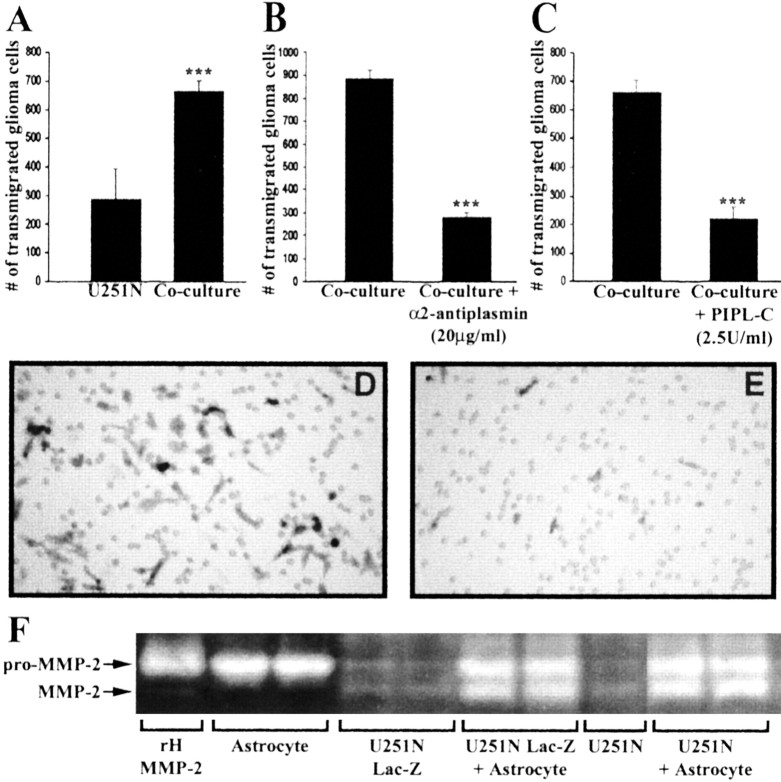
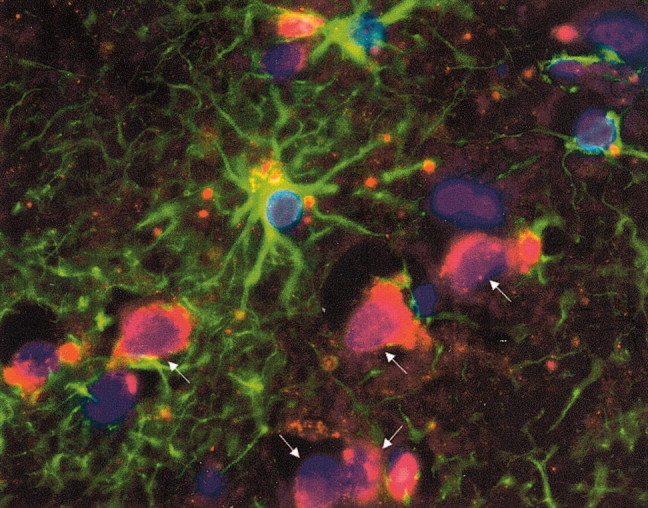
The presence of reactive astrocytes around glioma cells in the CNS suggests the possibility that these two cell types could be interacting. We addressed whether glioma cells use the astrocyte environment to modulate matrix metalloproteinase-2 (MMP-2), a proteolytic enzyme implicated in the invasiveness of glioma cells. We found that astrocytes in culture produce significant amounts of the pro-form of MMP-2 but undetectable levels of active MMP-2. However, after coculture with the U251N glioma line, astrocyte pro-MMP-2 was converted to the active form. The mechanism of pro-MMP-2 activation in glioma-astrocyte coculture was investigated and was found to involve the urokinase-type plasminogen activator (uPA)-plasmin cascade whereby uPA bound to uPA receptor (uPAR), leading to the conversion of plasminogen to plasmin. The latter cleaved pro-MMP-2 to generate its active form. Furthermore, key components (i.e., uPAR, uPA, and pro-MMP-2) were contributed principally by astrocytes, whereas the U251N glioma cells provided plasminogen. In correspondence with this biochemical cascade, the transmigration of U251N cells through Boyden invasion chambers coated with an extracellular matrix barrier was increased significantly in the presence of astrocytes, and this was inhibited by agents that disrupted the uPA-plasmin cascade. Finally, using resected human glioblastoma specimens, we found that tumor cells, but not astrocytes, expressed plasminogen in situ. We conclude that glioma cells exploit their astrocyte environment to activate MMP-2 and that this leads to the increased invasiveness of glioma cells.
Figures

References
-
- Apodaca G, Rutka JT, Bouhana K, Berens ME, Giblin JR, Rosenblum ML, McKerrow JH, Banda MJ ( 1990) Expression of metalloproteinases and metalloproteinase inhibitors by fetal astrocytes and glioma cells. Cancer Res 50: 2322–2329. - PubMed
-
- Beliveau R, Delbecchi L, Beaulieu E, Mousseau N, Kachra Z, Berthelet F, Moumdjian R, Del Maestro R ( 1999) Expression of matrix metalloproteinases and their inhibitors in human brain tumors. Ann NY Acad Sci 886: 236–239. - PubMed
-
- Bello L, Lucini V, Carrabba G, Giussani C, Machluf M, Pluderi M, Nikas D, Zhang J, Tomei G, Villani RM, Carroll RS, Bikfalvi A, Black PM ( 2001) Simultaneous inhibition of glioma angiogenesis, cell proliferation, and invasion by a naturally occurring fragment of human metalloproteinase-2. Cancer Res 61: 8730–8736. - PubMed
-
- Benbow U, Schoenermark MP, Mitchell TI, Rutter JL, Shimokawa K, Nagase H, Brinckerhoff CE ( 1999) A novel host/tumor cell interaction activates matrix metalloproteinase-1 and mediates invasion through type I collagen. J Biol Chem 274: 25371–25378. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous