

Expression and function of chloride transporters during development of inhibitory neurotransmission in the auditory brainstem
- PMID: 12764101
- PMCID: PMC6741087
- DOI: 10.1523/JNEUROSCI.23-10-04134.2003
Expression and function of chloride transporters during development of inhibitory neurotransmission in the auditory brainstem
Abstract
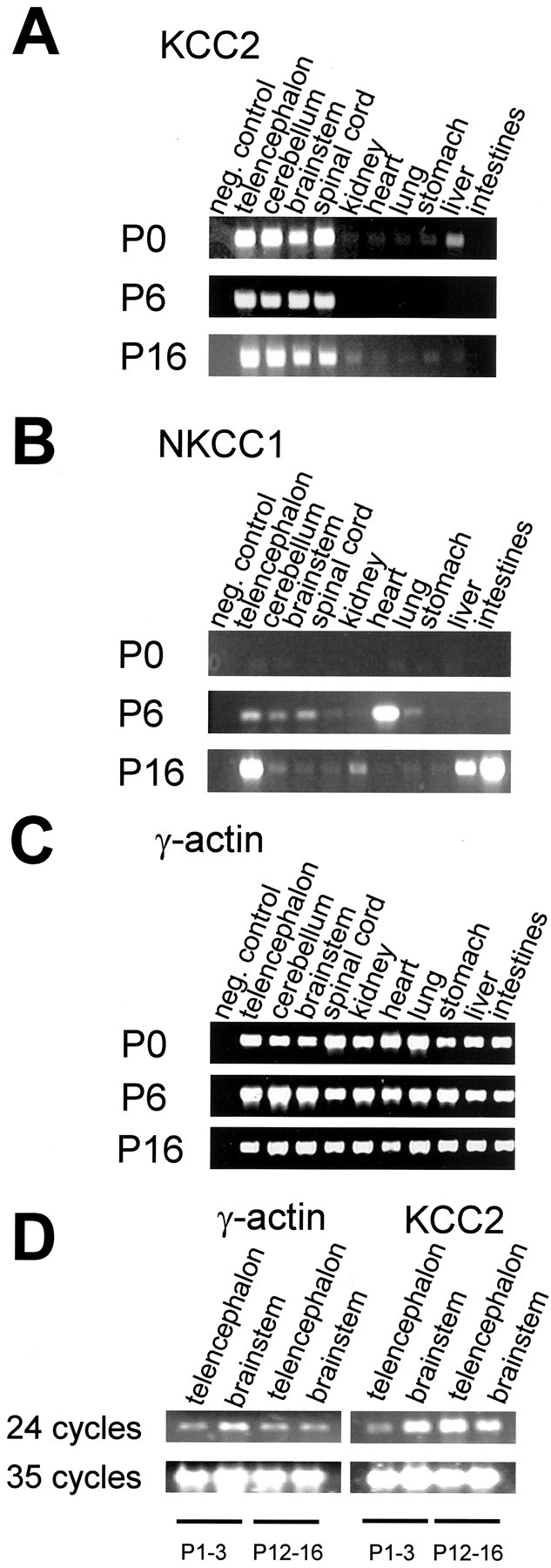
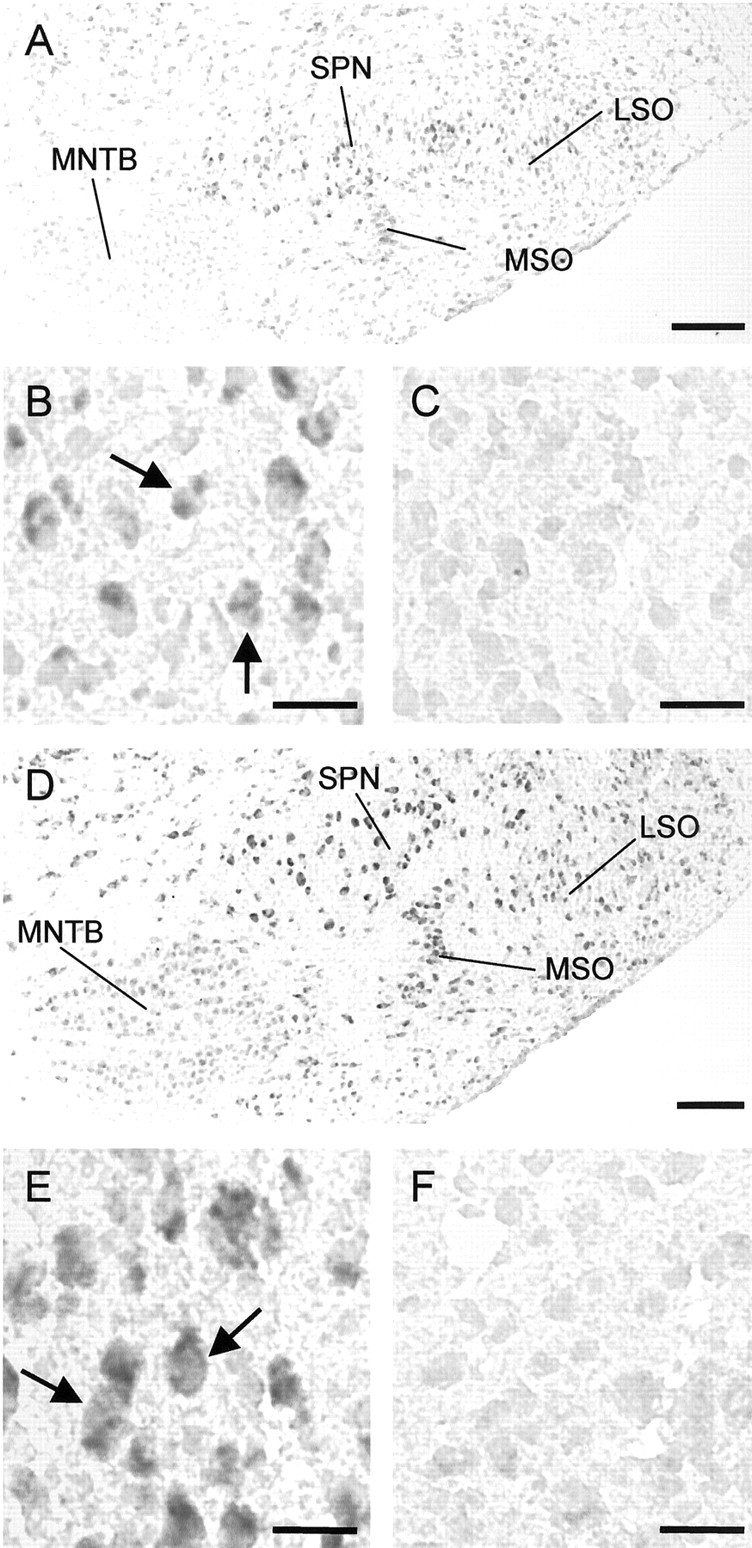
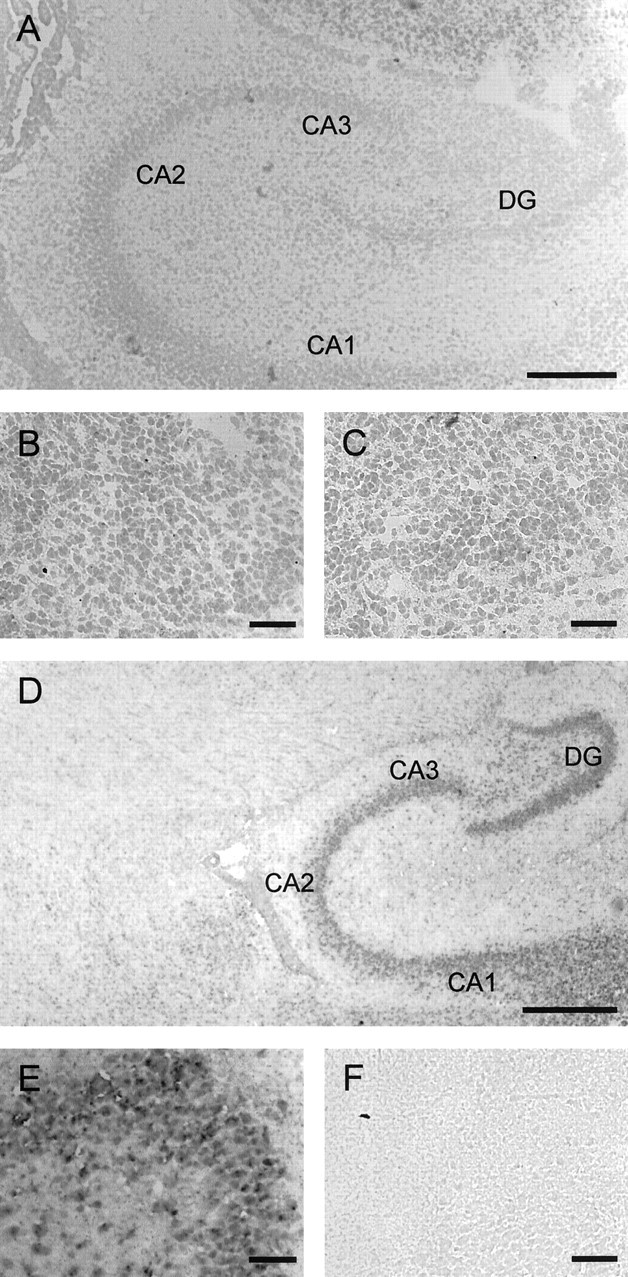
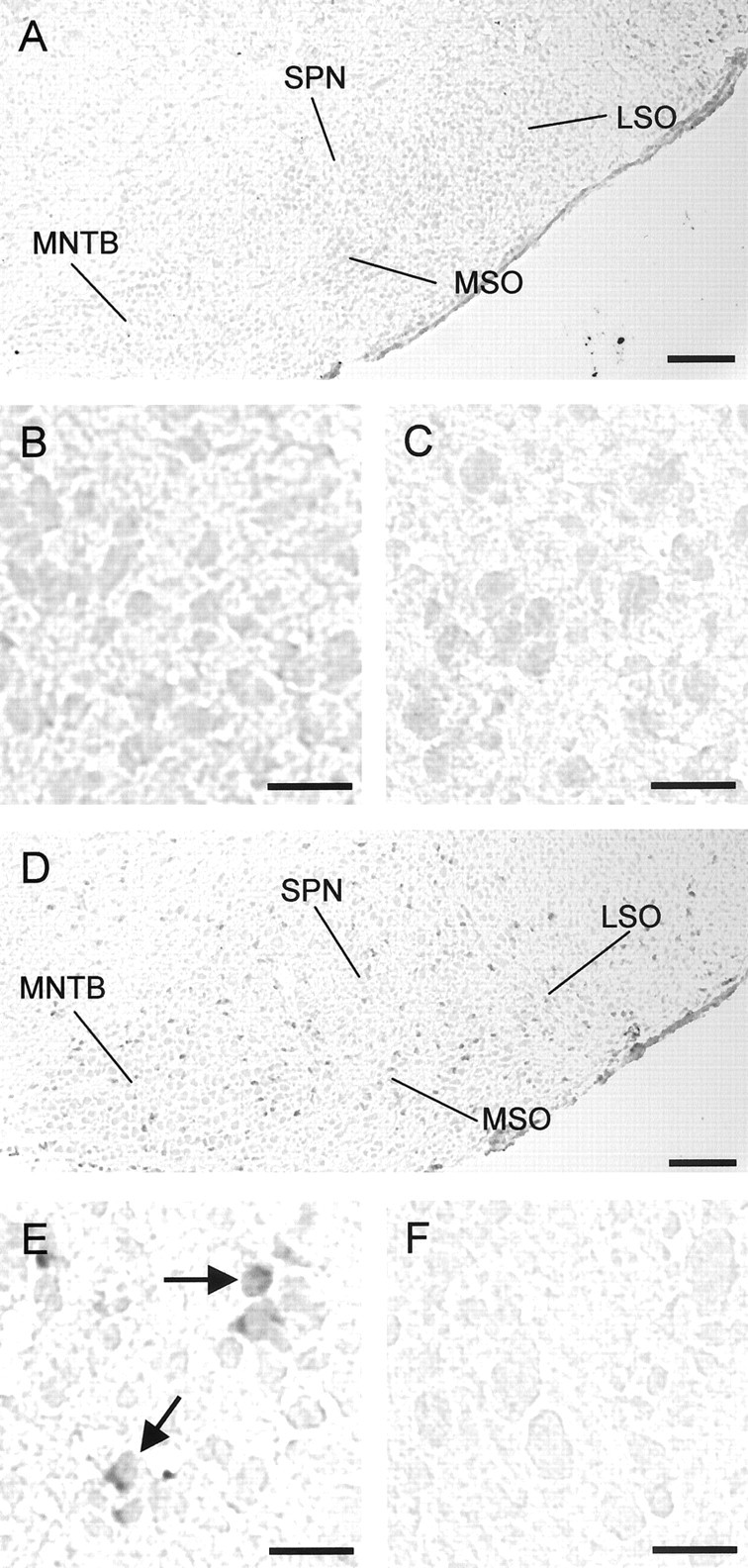
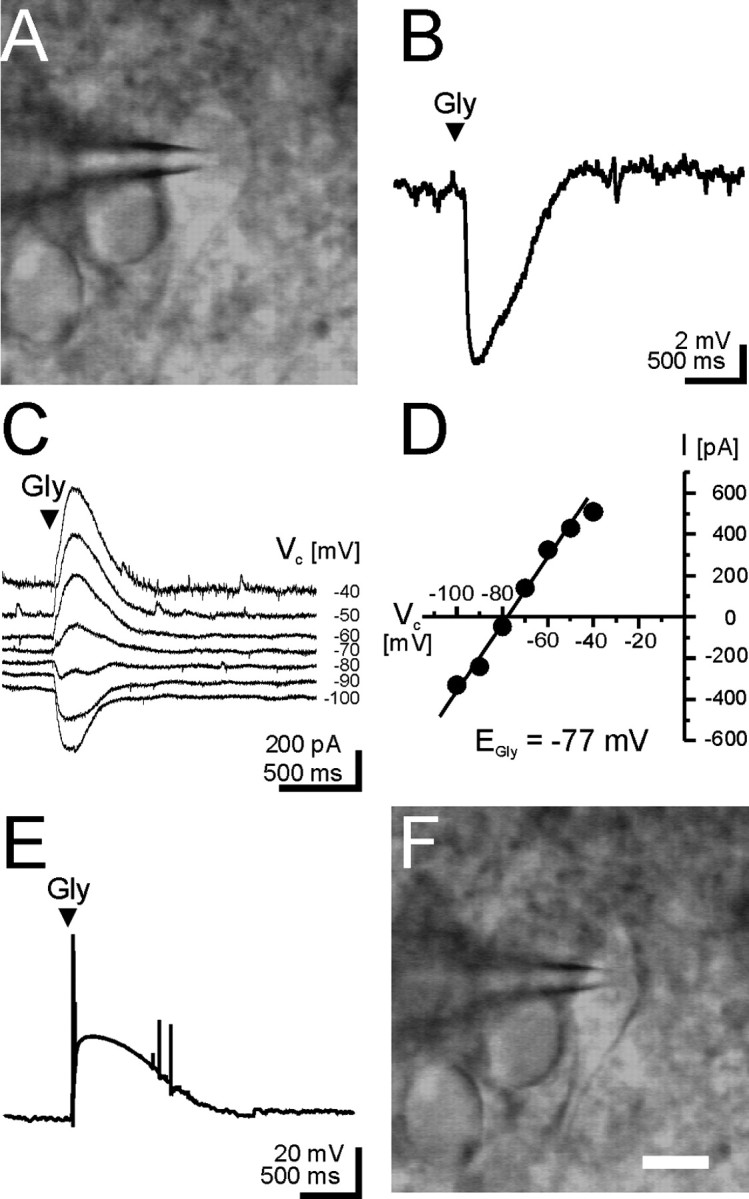
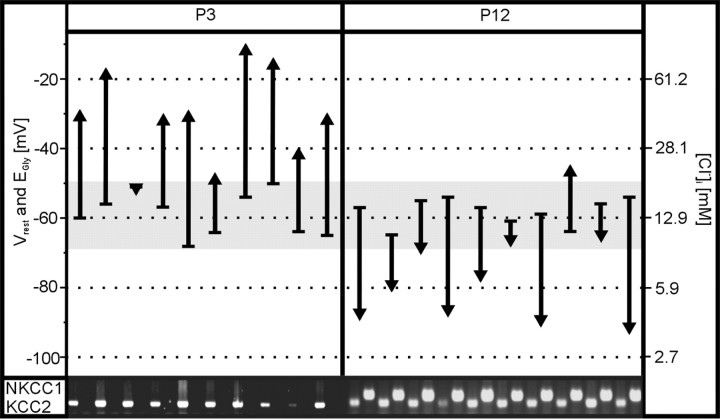
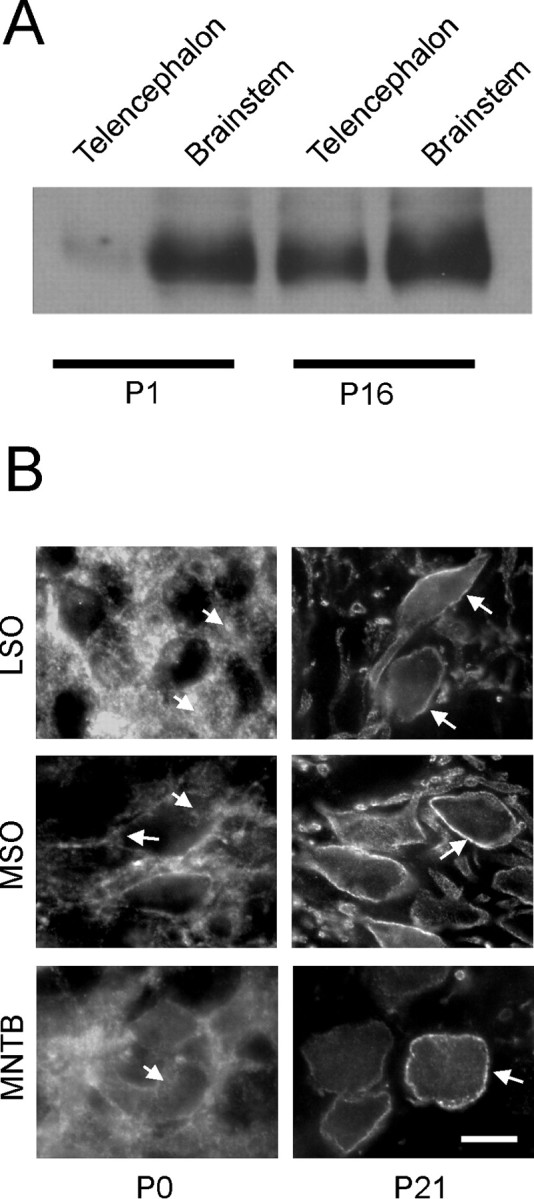
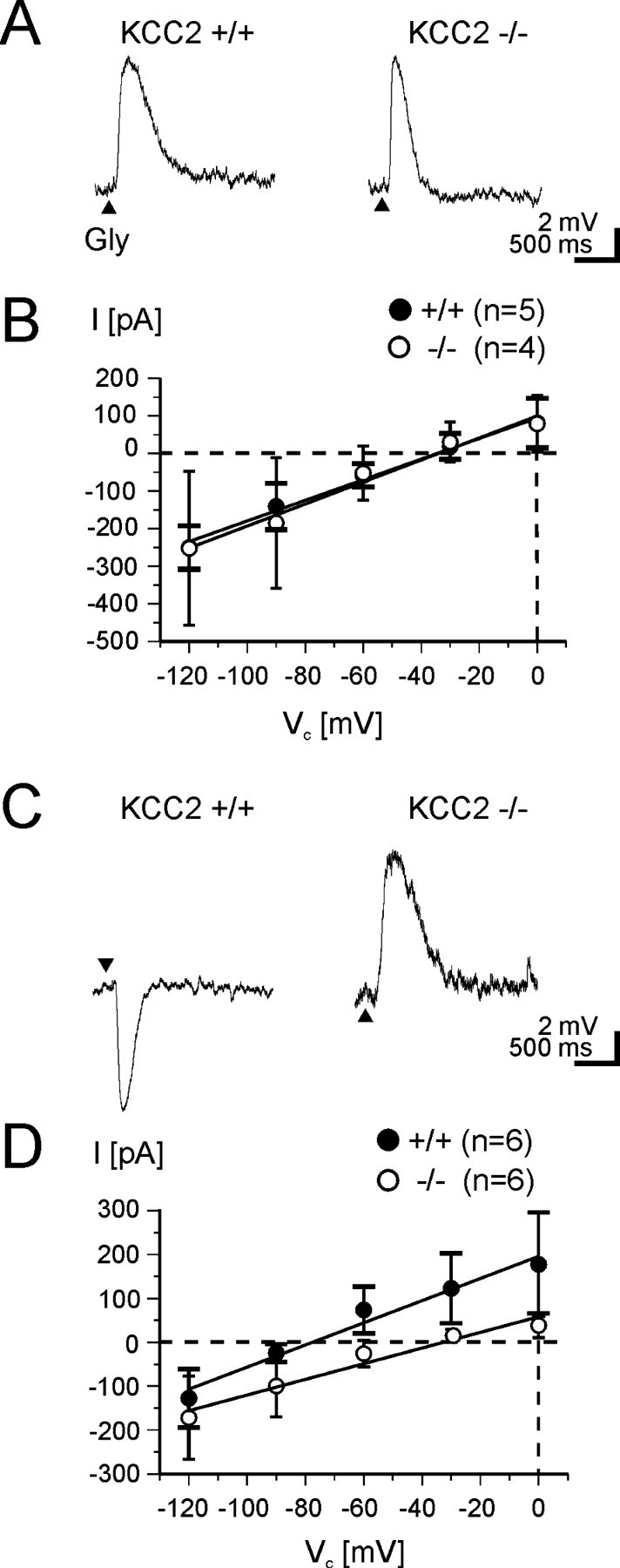
Glycine and GABA, the dominant inhibitory neurotransmitters in the CNS, assume a depolarizing role in early development, leading to increased cytoplasmic Ca2+ levels and action potentials. The effect is thought to be of some significance for maturation. The depolarization is caused by Cl- efflux, and chloride transporters contribute to the phenomenon by raising the intracellular Cl- concentration ([Cl-]i) above equilibrium, thereby generating an outward-directed electrochemical gradient for Cl-. In mature neurons, the [Cl-]i is reduced below equilibrium, thus rendering glycine activity hyperpolarizing. Here, we investigated the temporal expression of the K-Cl cotransporter KCC2 and the Na-K-Cl cotransporter NKCC1 in the lateral superior olive (LSO) of rats and mice. The two cation cotransporters normally extrude and accumulate Cl-, respectively. As evidenced by several methods, KCC2 mRNA was present in LSO neurons during both the depolarizing and hyperpolarizing periods. Western blots confirmed a constant level of KCC2 in the brainstem, and immunohistochemistry showed that the protein is diffusely distributed within neonatal LSO neurons, becoming integrated into the plasma membrane only with increasing age. The glycine reversal potential in KCC2 knock-out mice differed significantly from that determined in wild-type controls at postnatal day 12 (P12) but not at P3, demonstrating that KCC2 is not active in neonates, despite its early presence. NKCC1 mRNA was not detected during the depolarizing phase in the LSO, implying that this transporter does not contribute to the high [Cl-]i. Our results reveal major differences in the development of [Cl-]i regulation mechanisms seen in brainstem versus forebrain regions.
Figures
References
-
- Alvarez-Leefmans FJ ( 2001) Intracellular chloride regulation. In: Cell physiology sourcebook: a molecular approach (Sperelakis N, ed), pp 301–318. San Diego: Academic.
-
- Barker JL, Behar T, Li YX, Liu QY, Ma W, Maric D, Maric I, Schaffner AE, Serafini R, Smith SV, Somogyi R, Vautrin JY, Wen XL, Xian H ( 1998) GABAergic cells and signals in CNS development. Perspect Dev Neurobiol 5: 305–322. - PubMed
-
- Becker M, Nothwang HG, Friauf E ( 2003) Differential expression pattern of chloride transporters NCC, NKCC2, KCC1, KCC3, KCC4, and AE3 in the developing rat auditory brainstem. Cell Tiss Res, in press. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous