

Cloning of an avian adeno-associated virus (AAAV) and generation of recombinant AAAV particles
- PMID: 12768000
- PMCID: PMC156192
- DOI: 10.1128/jvi.77.12.6799-6810.2003
Cloning of an avian adeno-associated virus (AAAV) and generation of recombinant AAAV particles
Abstract
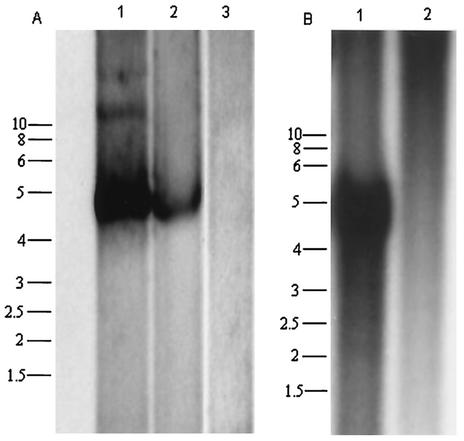
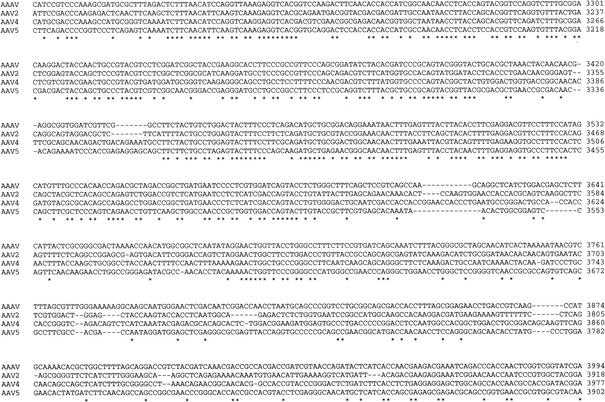
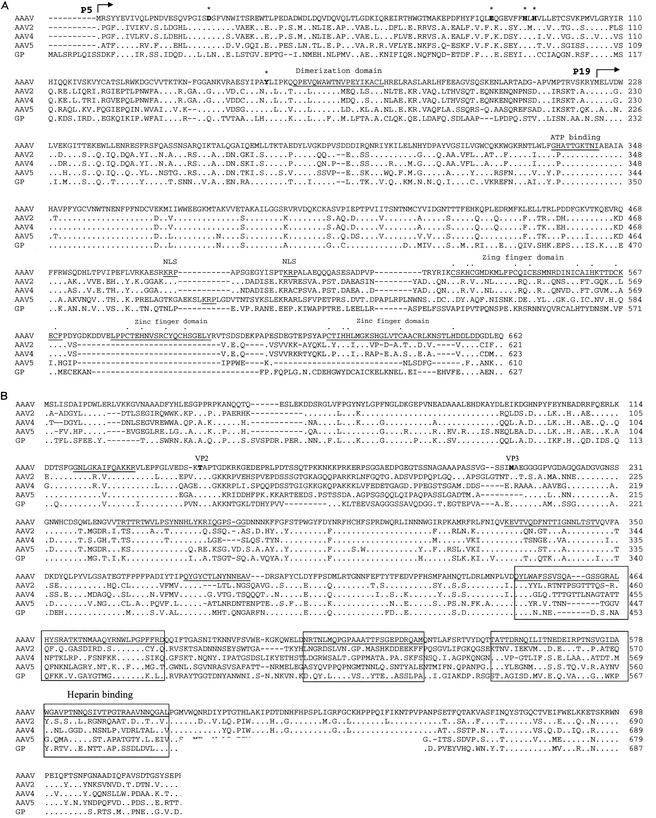
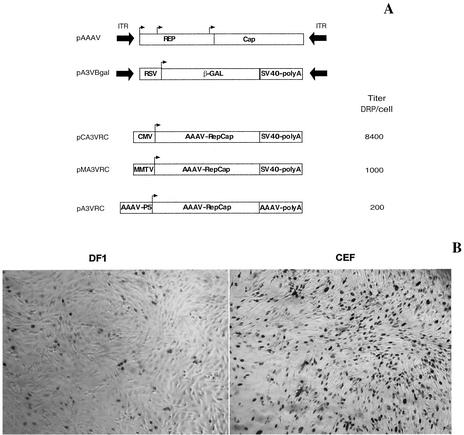
Recent studies have proposed that adeno-associated viruses (AAVs) are not evolutionarily linked to other mammalian autonomous parvoviruses but are more closely linked to the autonomous parvoviruses of birds. To better understand the relationship between primate and avian AAVs (AAAVs), we cloned and sequenced the genome of an AAAV (ATCC VR-865) and generated recombinant AAAV particles. The genome of AAAV is 4,694 nucleotides in length and has organization similar to that of other AAVs. The entire genome of AAAV displays 56 to 65% identity at the nucleotide level with the other known AAVs. The AAAV genome has inverted terminal repeats of 142 nucleotides, with the first 122 forming the characteristic T-shaped palindromic structure. The putative Rep-binding element consists of a tandem (GAGY)(4) repeat, and the putative terminal resolution site (trs), CCGGT/CG, contains a single nucleotide substitution relative to the AAV(2) trs. The Rep open reading frame of AAAV displays 50 to 54% identity at the amino acid level with the other AAVs, with most of the diversity clustered at the carboxyl and amino termini. Comparison of the capsid proteins of AAAV and the primate dependoviruses indicate that divergent regions are localized to surface-exposed loops. Despite these sequence differences, we were able to produce recombinant AAAV particles carrying a lacZ reporter gene by cotransfection in 293T cells and were able to examine transduction efficiency in both chicken primary cells and several cell lines. Our findings indicate that AAAV is the most divergent AAV described to date but maintains all the characteristics unique to the genera of dependovirus.
Figures






References
-
- Astell, C. R. 1990. Terminal hairpins of parvovirus genomes and their role in DNA replication, p. 59-79. In P. Tijssen (ed.), Handbook of parvoviruses. CRC Press, Boca Raton, Fla.
-
- Atchison, R. W., B. C. Casto, and W. Hammon. 1965. Adenovirus-associated defective virus particles. Science 149:754-756. - PubMed
-
- Bantel-Schall, U., and H. zur Hausen. 1984. Characterization of the DNA of a defective human parvovirus isolated from a genital site. Virology 134:52-63. - PubMed
-
- Bauer, H. J., and G. Monreal. 1986. Herpesviruses provide helper functions for avian adeno-associated parvovirus. J. Gen. Virol. 67:181-185. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
