

Interactions of rotavirus VP4 spike protein with the endosomal protein Rab5 and the prenylated Rab acceptor PRA1
- PMID: 12768023
- PMCID: PMC156175
- DOI: 10.1128/jvi.77.12.7041-7047.2003
Interactions of rotavirus VP4 spike protein with the endosomal protein Rab5 and the prenylated Rab acceptor PRA1
Abstract
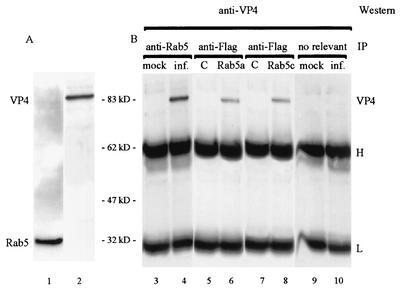
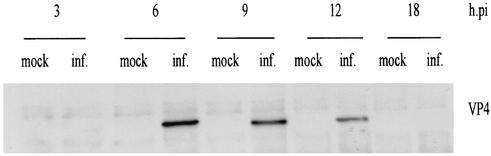
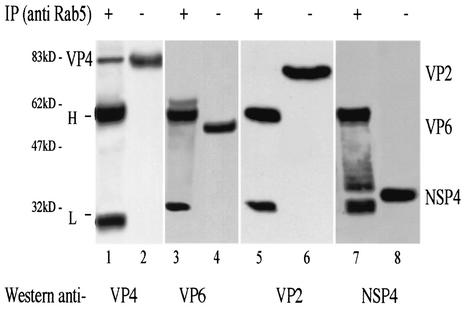
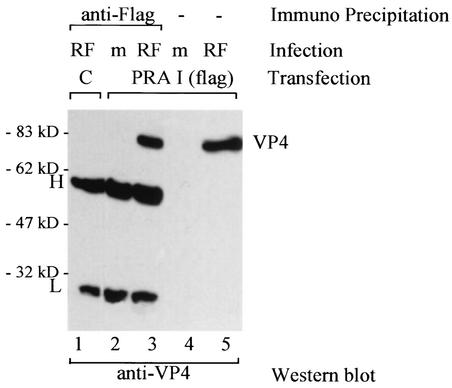
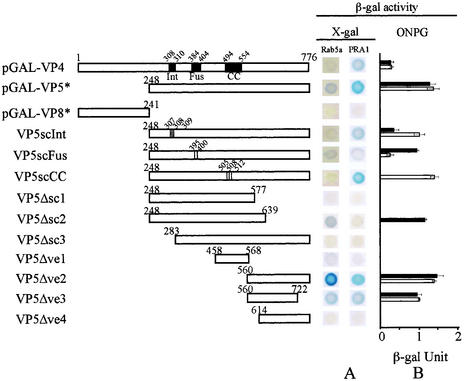
Rotavirus spike protein VP4 is implicated in several important functions, such as cell attachment, penetration, hemagglutination, neutralization, virulence, and host range. It is present at the plasma membrane and colocalizes with the cytoskeleton in infected cells. We looked for cellular partners responsible for the localization of VP4 by two-hybrid screening of a monkey CV1 cell cDNA library. In the screen we isolated repeatedly three cDNAs encoding either two isoforms (a and c) of Rab5 protein or the prenylated Rab acceptor (PRA1). The small GTPase Rab5 is a molecule regulating the vesicular traffic and the motility of early endosomes along microtubules. Rab5 interacts with a large number of effectors, in particular with PRA1. Interactions of VP4 with both partners, Rab5 and PRA1, were confirmed by coimmunoprecipitation from infected- or transfected-cell lysates. Interaction of Rab5 and PRA1 was restricted to free VP4, since neither triple-layered particles nor NSP4-VP4-VP7 heterotrimeric complexes could be coprecipitated. Site-directed and deletion mutants of VP4 were used to map a VP4 domain(s) interacting with Rab5 or PRA1. Of the 10 mutants tested, 2 interacted exclusively with a single partner. In contrast, the domain extending from amino acids 560 to 722 of VP4 is essential for both interactions. These results suggest that Rab5 and PRA1 may be involved in the localization and trafficking of VP4 in infected cells.
Figures
References
-
- Abdul-Ghani, M., P. Y. Gougeon, D. C. Prosser, L. F. Da-Silva, and J. K. Ngsee. 2001. PRA isoforms are targeted to distinct membrane compartments. J. Biol. Chem. 276:6225-6233. - PubMed
-
- Belin, M. T., and P. Boulanger. 1993. Involvement of cellular adhesion sequences in the attachment of adenovirus to the HeLa cell surface. J. Gen. Virol. 74:1485-1497. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
