

In vitro analysis of nuclear mRNA export using molecular beacons for target detection
- PMID: 12771228
- PMCID: PMC156741
- DOI: 10.1093/nar/gng063
In vitro analysis of nuclear mRNA export using molecular beacons for target detection
Abstract
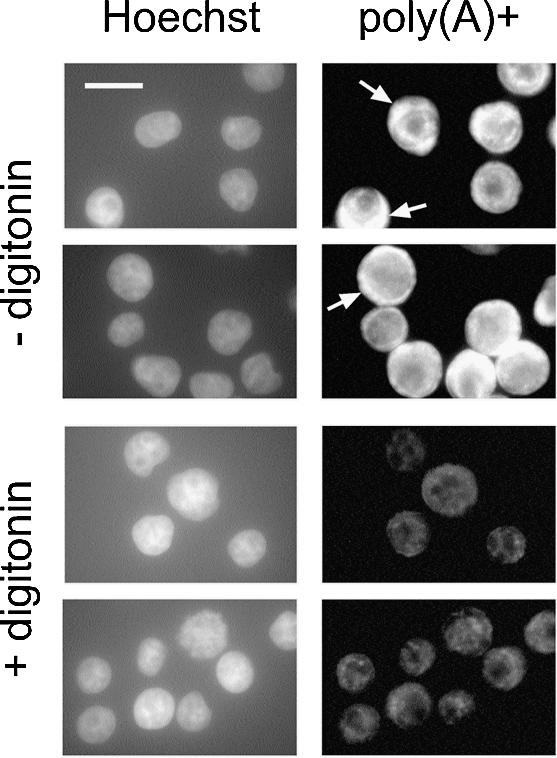
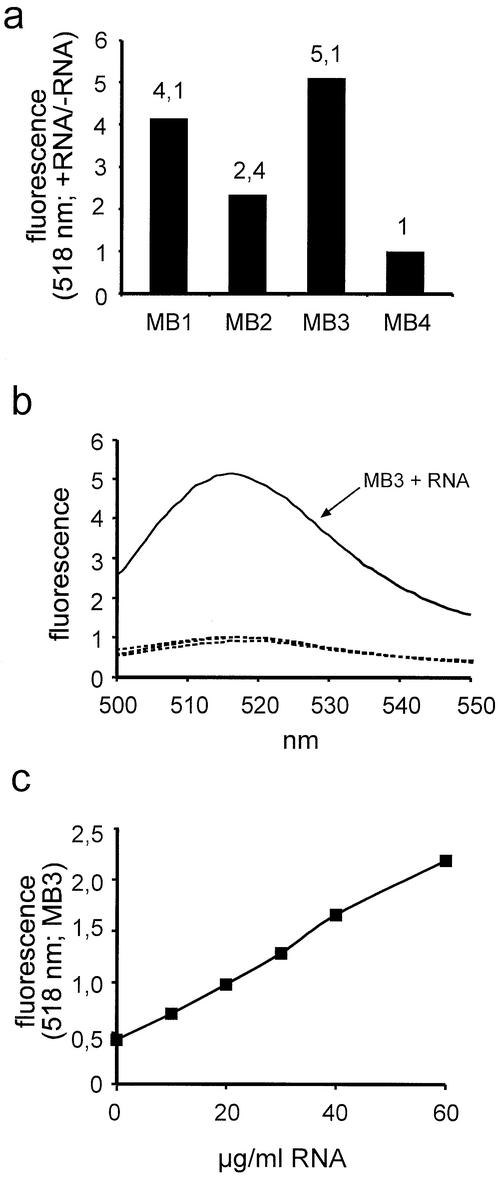
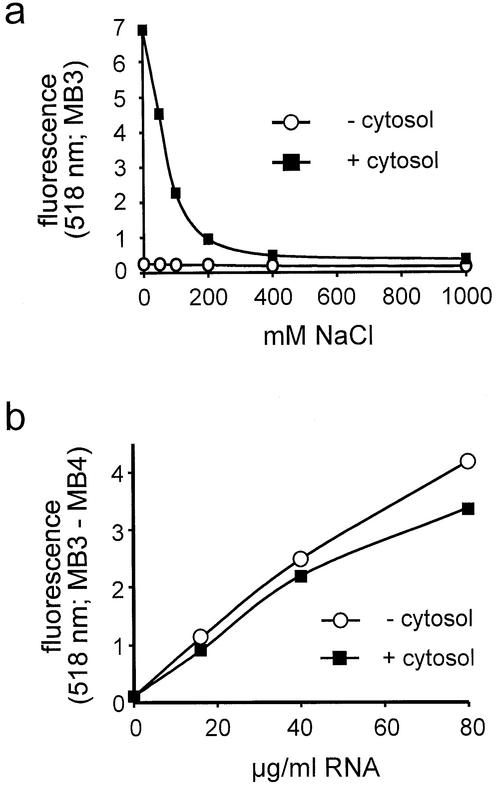
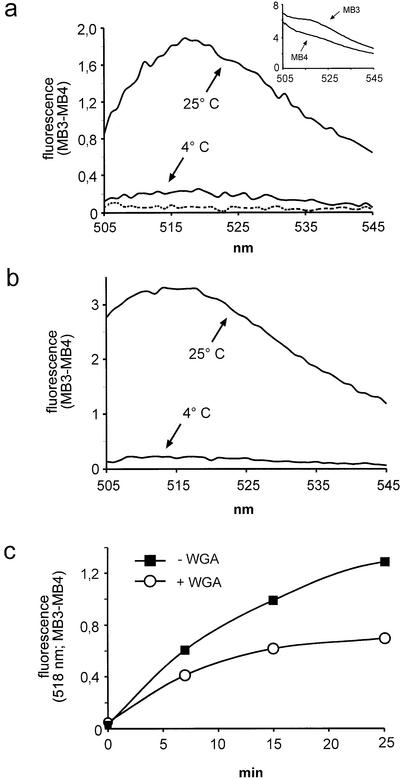
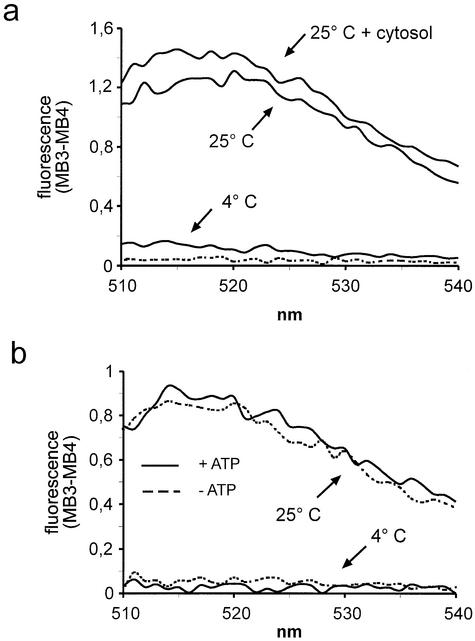
A detailed molecular characterization of nuclear mRNA export will require an in vitro system, allowing a biochemical reconstitution of transport. To this end, an mRNA export assay has been developed using digitonin-permeabilized HeLa cells and 2'-O-methyl oligoribonucleotide molecular beacons for target detection. These probes allow the homogeneous detection of poly(A)+ RNA at subnanomolar concentrations in the presence of cytosol, without the need for RNA purification and time-consuming methods like northern blotting or RT-PCR. Nuclear export of endogenous mRNA in permeabilized cells occurs in a time- and temperature-dependent manner and can be inhibited by wheat germ agglutinin, indicative of specific transport through nuclear pore complexes. Nuclear export in vitro is insensitive to the depletion of ATP and does not depend on the addition of cytosolic factors, suggesting that shuttling proteins are not required for efficient transport. This is the first demonstration of molecular beacons as powerful tools for the analysis of nucleocytoplasmic RNA transport.
Figures
References
-
- Görlich D. and Kutay,U. (1999) Transport between the cell nucleus and the cytoplasm. Annu. Rev. Cell. Dev. Biol., 15, 607–660. - PubMed
-
- Weis K. (2003) Regulating access to the genome: nucleocytoplasmic transport throughout the cell cycle. Cell, 112, 441–451. - PubMed
-
- Fahrenkrog B., Stoffler,D. and Aebi,U. (2001) Nuclear pore complex architecture and functional dynamics. Curr. Top. Microbiol. Immunol., 259, 95–117. - PubMed
-
- Kalab P., Weis,K. and Heald,R. (2002) Visualization of a Ran-GTP gradient in interphase and mitotic Xenopus egg extracts. Science, 295, 2452–2456. - PubMed
