

Structure of Rhodococcus erythropolis limonene-1,2-epoxide hydrolase reveals a novel active site
- PMID: 12773375
- PMCID: PMC156771
- DOI: 10.1093/emboj/cdg275
Structure of Rhodococcus erythropolis limonene-1,2-epoxide hydrolase reveals a novel active site
Abstract
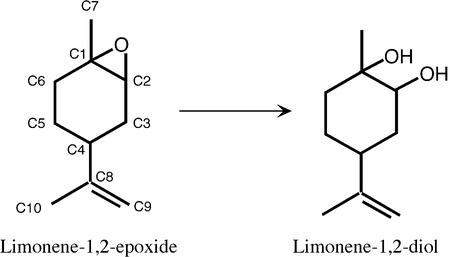
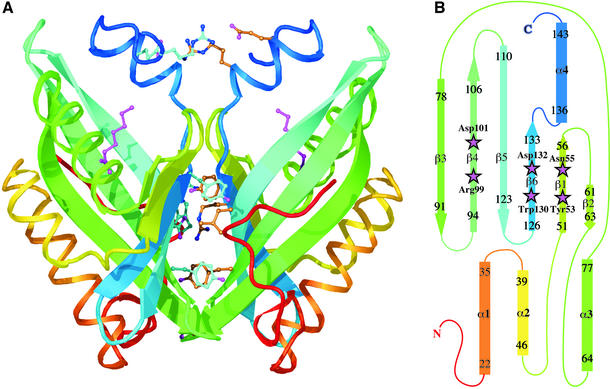
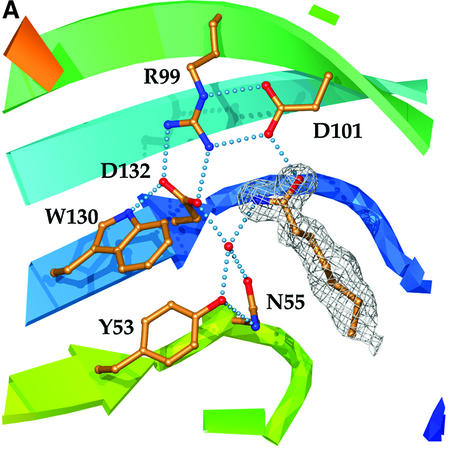
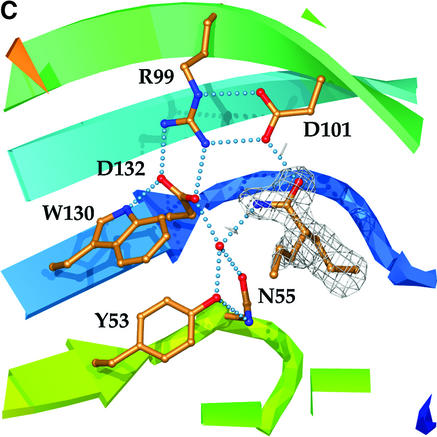
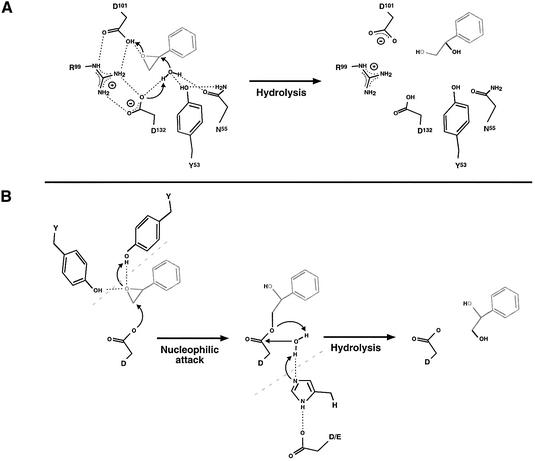
Epoxide hydrolases are essential for the processing of epoxide-containing compounds in detoxification or metabolism. The classic epoxide hydrolases have an alpha/beta hydrolase fold and act via a two-step reaction mechanism including an enzyme-substrate intermediate. We report here the structure of the limonene-1,2-epoxide hydrolase from Rhodococcus erythropolis, solved using single-wavelength anomalous dispersion from a selenomethionine-substituted protein and refined at 1.2 A resolution. This enzyme represents a completely different structure and a novel one-step mechanism. The fold features a highly curved six-stranded mixed beta-sheet, with four alpha-helices packed onto it to create a deep pocket. Although most residues lining this pocket are hydrophobic, a cluster of polar groups, including an Asp-Arg-Asp triad, interact at its deepest point. Site-directed mutagenesis supports the conclusion that this is the active site. Further, a 1.7 A resolution structure shows the inhibitor valpromide bound at this position, with its polar atoms interacting directly with the residues of the triad. We suggest that several bacterial proteins of currently unknown function will share this structure and, in some cases, catalytic properties.
Figures


References
-
- Archelas A. and Furstoss,R. (2001) Synthetic applications of epoxide hydrolases. Curr. Opin. Chem. Biol., 5, 112–119. - PubMed
-
- Argiriadi M.A., Morisseau,C., Goodrow,M.H., Dowdy,D.L., Hammock,B.D. and Christianson,D.W. (2000) Binding of alkylurea inhibitors to epoxide hydrolase implicates active site tyrosines in substrate activation. J. Biol. Chem., 275, 15265–15270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
