

The role of Ppe1/PP6 phosphatase for equal chromosome segregation in fission yeast kinetochore
- PMID: 12773390
- PMCID: PMC156763
- DOI: 10.1093/emboj/cdg266
The role of Ppe1/PP6 phosphatase for equal chromosome segregation in fission yeast kinetochore
Abstract
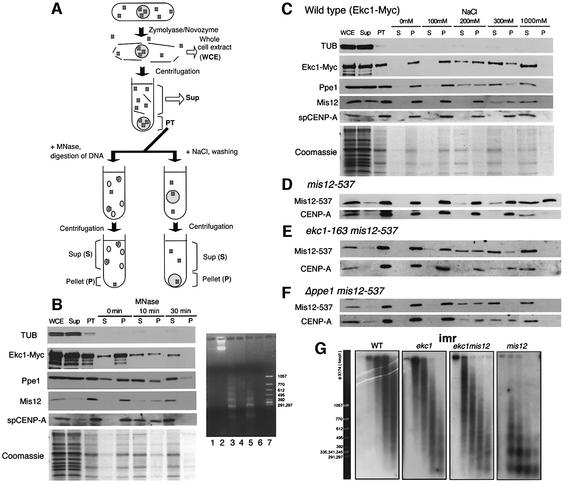
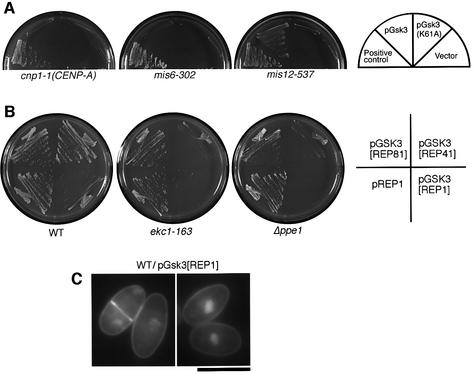
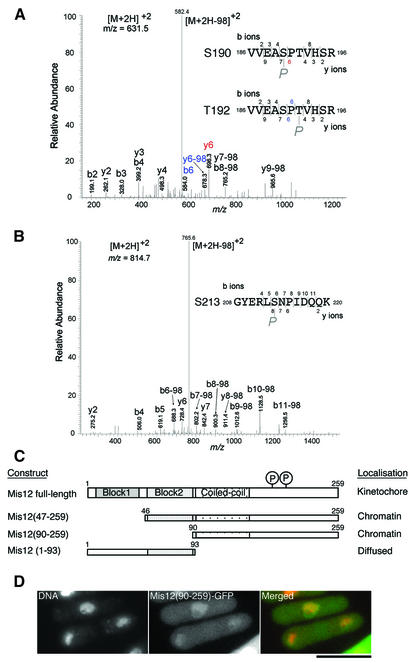
Mis12 is a kinetochore protein essential for equal chromosome segregation and is evolutionarily conserved from yeast to human. In this study, we report the isolation and characterization of suppressors of the mis12 mutant in fission yeast. Our results indicate that Mis12 is negatively regulated by a highly conserved protein phosphatase Ppe1 (scSit4/dmPPV/hPP6) or its bound partner Ekc1 (scSAP), and it is positively regulated by a counteracting kinase Gsk3. Mass spectrometry analysis shows that at least two sites in Mis12 are phosphorylated. This mechanism of suppression occurs at the level of localization recovery of Mis12 to the kinetochore chromatin. Consistently, Mis12 and a subpopulation of Ppe1/Ekc1 were found to behave like non-histone-type chromatin-associating proteins in the chromatin fractionation assay. Mutant analysis of Ppe1 and Ekc1 revealed that they are important for faithful chromosome segregation, as the mutants exhibited unequal chromosome segregation similar to mis12 in the presence of a low concentration of tubulin poison. Ppe1/PP6 directly or indirectly modulates kinetochore chromatin protein Mis12 to ensure progression into normal anaphase.
Figures
Similar articles
-
CDC2 phosphorylation of the fission yeast dis1 ensures accurate chromosome segregation.Curr Biol. 2006 Aug 22;16(16):1627-35. doi: 10.1016/j.cub.2006.06.065. Curr Biol. 2006. PMID: 16920624
-
A conserved Mis12 centromere complex is linked to heterochromatic HP1 and outer kinetochore protein Zwint-1.Nat Cell Biol. 2004 Nov;6(11):1135-41. doi: 10.1038/ncb1187. Epub 2004 Oct 24. Nat Cell Biol. 2004. PMID: 15502821
-
Schizosaccharomyces pombe cell division cycle under limited glucose requires Ssp1 kinase, the putative CaMKK, and Sds23, a PP2A-related phosphatase inhibitor.Genes Cells. 2009 May;14(5):539-54. doi: 10.1111/j.1365-2443.2009.01290.x. Epub 2009 Apr 15. Genes Cells. 2009. PMID: 19371376
-
Two distinct pathways responsible for the loading of CENP-A to centromeres in the fission yeast cell cycle.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):595-606; discussion 606-7. doi: 10.1098/rstb.2004.1614. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897182 Free PMC article. Review.
-
[Kinetochore geometry defined by cohesin within the centromere].Tanpakushitsu Kakusan Koso. 2009 Nov;54(14):1842-9. Tanpakushitsu Kakusan Koso. 2009. PMID: 19894585 Review. Japanese. No abstract available.
Cited by
-
Genetic Interactions among AMPK Catalytic Subunit Ssp2 and Glycogen Synthase Kinases Gsk3 and Gsk31 in Schizosaccharomyces Pombe.Kobe J Med Sci. 2016 Aug 3;62(3):E70-8. Kobe J Med Sci. 2016. PMID: 27604537 Free PMC article.
-
Negative Regulation of the Mis17-Mis6 Centromere Complex by mRNA Decay Pathway and EKC/KEOPS Complex in Schizosaccharomyces pombe.G3 (Bethesda). 2019 Jun 5;9(6):1815-1823. doi: 10.1534/g3.119.400227. G3 (Bethesda). 2019. PMID: 30967422 Free PMC article.
-
Isolation of Fission Yeast Condensin Temperature-Sensitive Mutants with Single Amino Acid Substitutions Targeted to Hinge Domain.G3 (Bethesda). 2019 May 7;9(5):1777-1783. doi: 10.1534/g3.119.400156. G3 (Bethesda). 2019. PMID: 30914423 Free PMC article.
-
Genetic-interaction screens uncover novel biological roles and regulators of transcription factors in fission yeast.G3 (Bethesda). 2022 Aug 25;12(9):jkac194. doi: 10.1093/g3journal/jkac194. G3 (Bethesda). 2022. PMID: 35924983 Free PMC article.
-
Whole-Genome Sequencing of Suppressor DNA Mixtures Identifies Pathways That Compensate for Chromosome Segregation Defects in Schizosaccharomyces pombe.G3 (Bethesda). 2018 Mar 2;8(3):1031-1038. doi: 10.1534/g3.118.200048. G3 (Bethesda). 2018. PMID: 29352077 Free PMC article.
References
-
- Bastians H. and Ponstingl,H. (1996) The novel human protein serine/threonine phosphatase 6 is a functional homologue of budding yeast Sit4p and fission yeast ppe1, which are involved in cell cycle regulation. J. Cell Sci., 109, 2865–2874. - PubMed
-
- Beck T. and Hall,M.N. (1999) The TOR signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature, 402, 689–692. - PubMed
-
- Cohen P.T. (1997) Novel protein serine/threonine phosphatases: variety is the spice of life. Trends Biochem. Sci., 22, 245–251. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
