

Cysteine 64 of Ref-1 is not essential for redox regulation of AP-1 DNA binding
- PMID: 12773568
- PMCID: PMC156143
- DOI: 10.1128/MCB.23.12.4257-4266.2003
Cysteine 64 of Ref-1 is not essential for redox regulation of AP-1 DNA binding
Abstract
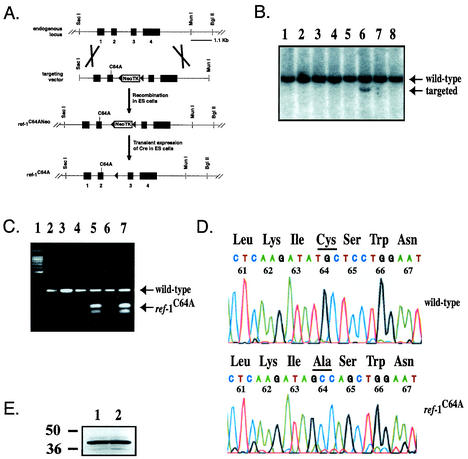
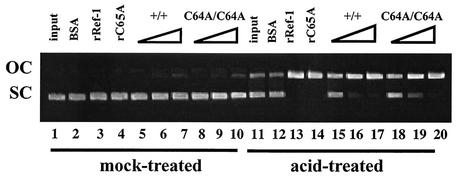
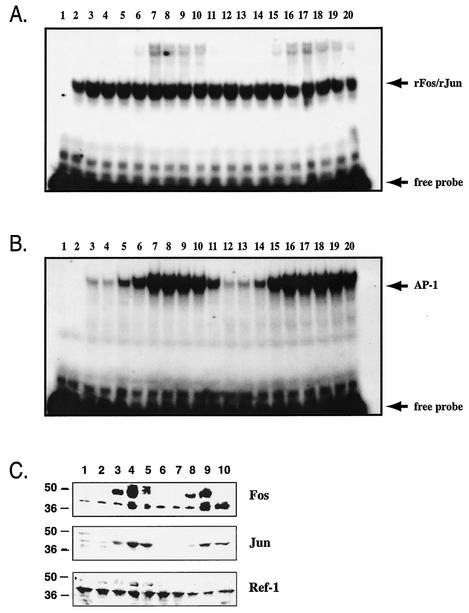
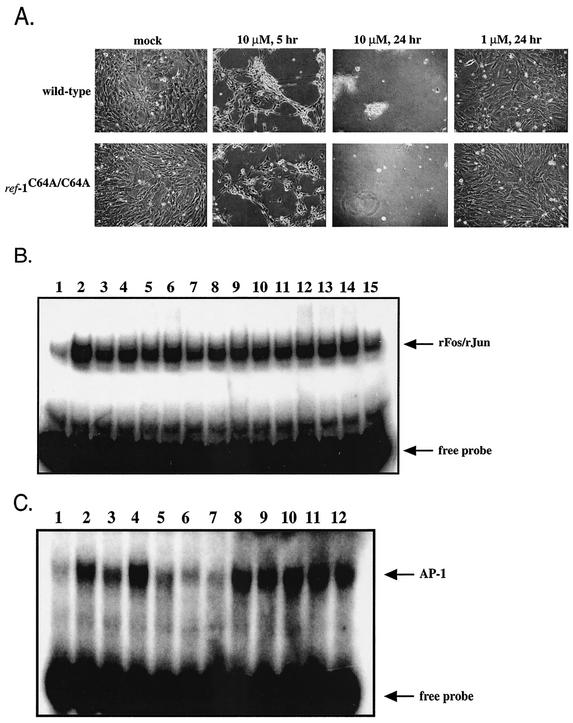
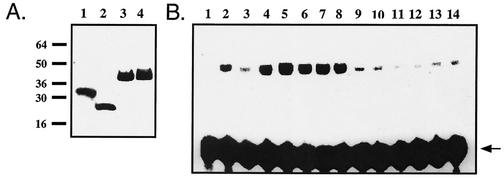
Ref-1 participates in DNA repair as well as in redox regulation of transcription factor function. The redox function of Ref-1 involves reduction of oxidized cysteine residues within the DNA binding domains of several transcription factors, including Fos and Jun. Reduction of these residues is required for DNA binding, providing a redox-dependent mechanism for regulation of target gene expression. Previous in vitro studies implicated cysteine 65 of human Ref-1 (cysteine 64 of mouse Ref-1) as the redox catalytic site. We analyzed the in vivo role of cysteine 64 in redox regulation of AP-1 activity by introducing a cysteine-to-alanine point mutation into the endogenous mouse Ref-1 gene (ref-1(C64A)). Unlike Ref-1 null mice, which die very early in embryonic development, homozygous ref-1(C64A) mice are viable, they survive to normal life expectancy, and they display no overt abnormal phenotype. Although Ref-1 provides the major AP-1-reducing activity in murine cells, ref-1(C64A) cells retain normal levels of endogenous AP-1 DNA binding activity in vivo as well as normal Fos- and Jun-reducing activity in vitro. These results demonstrate that Ref-1 cysteine 64/65 is not required for redox regulation of AP-1 DNA binding in vivo, and they challenge previous hypotheses regarding the mechanism by which Ref-1 regulates the redox-dependent activity of specific transcription factors.
Figures
References
-
- Abate, C., L. Patel, F. J. Rauscher III, and T. Curran. 1990. Redox regulation of fos and jun DNA-binding activity in vitro. Science 249:1157-1161. - PubMed
-
- Beiqing, L., M. Chen, and R. L. Whisler. 1996. Sublethal levels of oxidative stress stimulate transcriptional activation of c-jun and suppress IL-2 promoter activation in Jurkat T cells. J. Immunol. 157:160-169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous