

Cu/Zn superoxide dismutase mutants associated with amyotrophic lateral sclerosis show enhanced formation of aggregates in vitro
- PMID: 12773627
- PMCID: PMC165823
- DOI: 10.1073/pnas.1237797100
Cu/Zn superoxide dismutase mutants associated with amyotrophic lateral sclerosis show enhanced formation of aggregates in vitro
Abstract
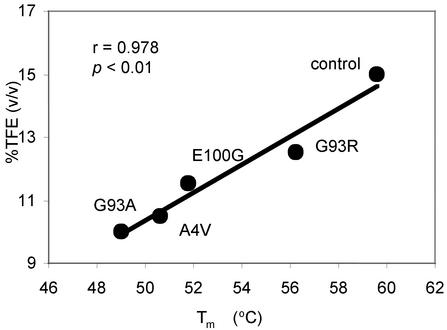
Mutations in Cu/Zn superoxide dismutase (SOD) are associated with the fatal neurodegenerative disorder amyotrophic lateral sclerosis (ALS). There is considerable evidence that mutant SOD has a gain of toxic function; however, the mechanism of this toxicity is not known. We report here that purified SOD forms aggregates in vitro under destabilizing solution conditions by a process involving a transition from small amorphous species to fibrils. The assembly process and the tinctorial and structural properties of the in vitro aggregates resemble those for aggregates observed in vivo. Furthermore, the familial ALS SOD mutations A4V, G93A, G93R, and E100G decrease protein stability, which correlates with an increase in the propensity of the mutants to form aggregates. These mutations also increase the rate of protein unfolding. Our results suggest three possible mechanisms for the increase in aggregation: (i) an increase in the equilibrium population of unfolded or of partially unfolded states, (ii) an increase in the rate of unfolding, and (iii) a decrease in the rate of folding. Our data support the hypothesis that the gain of toxic function for many different familial ALS-associated mutant SODs is a consequence of protein destabilization, which leads to an increase in the formation of cytotoxic protein aggregates.
Figures





References
-
- Fridovich, I. (1986) Adv. Enzymol. Relat. Areas Mol. Biol. 58, 61–97. - PubMed
-
- Lepock, J. R., Frey, H. E. & Hallewell, R. A. (1990) J. Biol. Chem. 265, 21612–21618. - PubMed
-
- Rosen, D. R., Siddique, T., Patterson, D., Figlewicz, D. A., Sapp, P., Hentati, A., Donaldson, D., Goto, J., O'Regan, J. P., Deng, H. X., et al. (1993) Nature 362, 59–62. - PubMed
-
- Rowland, L. P. & Shneider, N. A. (2001) N. Engl. J. Med. 344, 1688–1700. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
