

Protection and compensation in the influenza virus-specific CD8+ T cell response
- PMID: 12775762
- PMCID: PMC165859
- DOI: 10.1073/pnas.1232449100
Protection and compensation in the influenza virus-specific CD8+ T cell response
Abstract
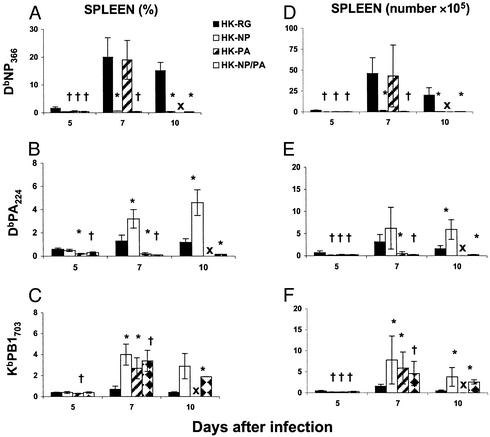
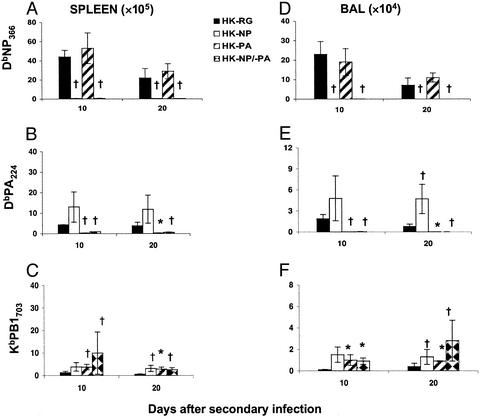
Influenza virus-specific CD8+ T cells generally recognize peptides derived from conserved, internal proteins that are not subject to antibody-mediated selection pressure. Prior exposure to any one influenza A virus (H1N1) can prime for a secondary CD8+ T cell response to a serologically different influenza A virus (H3N2). The protection afforded by this recall of established CD8+ T cell memory, although limited, is not negligible. Key characteristics of primary and secondary influenza-specific host responses are probed here with recombinant viruses expressing modified nucleoprotein (NP) and acid polymerase (PA) genes. Point mutations were introduced into the epitopes derived from the NP and PA such that they no longer bound the presenting H2Db MHC class I glycoprotein, and reassortant H1N1 and H3N2 viruses were made by reverse genetics. Conventional (C57BL/6J, H2b, and Ig+/+) and Ig-/- (muMT) mice were more susceptible to challenge with the single NP [HKx31 influenza A virus (HK)-NP] and PA (HK-PA) mutants, but unlike the Ig-/- mice, Ig+/+ mice were surprisingly resistant to the HK-NP/-PA double mutant. This virus was found to promote an enhanced IgG response resulting, perhaps, from the delayed elimination of antigen-presenting cells. Antigen persistence also could explain the increase in size of the minor KbPB1703 CD8+ T cell population in mice infected with the mutant viruses. The extent of such compensation was always partial, giving the impression that any virus-specific CD8+ T cell response operates within constrained limits. It seems that the relationship between protective humoral and cellular immunity is neither simple nor readily predicted.
Figures



References
-
- Yewdell, J. W. & Bennink, J. R. (1999) Annu. Rev. Immunol. 17, 51-88. - PubMed
-
- Belz, G. T., Xie, W. & Doherty, P. C. (2001) J. Immunol. 166, 4627-4633. - PubMed
-
- Townsend, A. R., Rothbard, J., Gotch, F. M., Bahadur, G., Wraith, D. & McMichael, A. J. (1986) Cell 44, 959-968. - PubMed
-
- Falk, K., Rotzschke, O., Stevanovic, S., Jung, G. & Rammensee, H.-G. (1991) Nature 351, 290-296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
