

Lateral gene transfer and the evolution of plastid-targeted proteins in the secondary plastid-containing alga Bigelowiella natans
- PMID: 12777624
- PMCID: PMC164647
- DOI: 10.1073/pnas.1230951100
Lateral gene transfer and the evolution of plastid-targeted proteins in the secondary plastid-containing alga Bigelowiella natans
Abstract
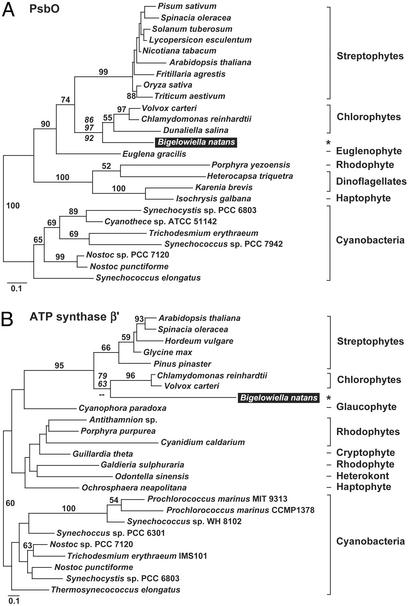
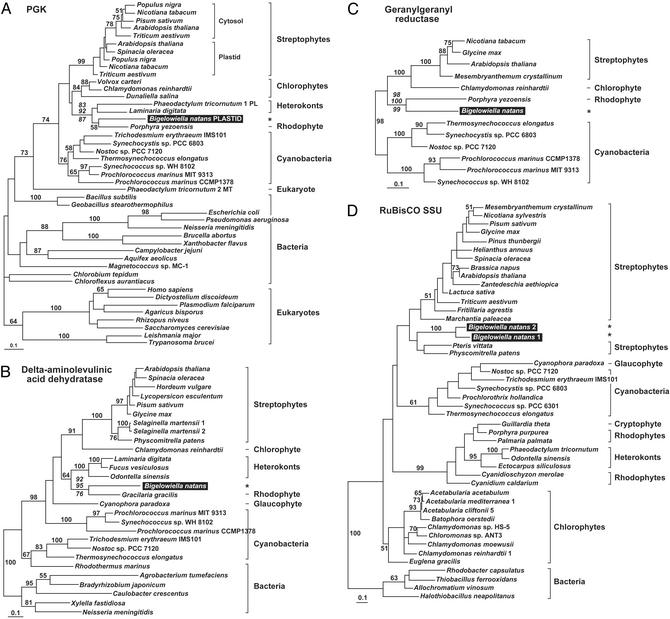
Chlorarachniophytes are amoeboflagellate algae that acquired photosynthesis secondarily by engulfing a green alga and retaining its plastid (chloroplast). An important consequence of secondary endosymbiosis in chlorarachniophytes is that most of the nuclear genes encoding plastid-targeted proteins have moved from the nucleus of the endosymbiont to the host nucleus. We have sequenced and analyzed 83 cDNAs encoding 78 plastid-targeted proteins from the model chlorarachniophyte Bigelowiella natans (formerly Chlorarachnion sp. CCMP621). Phylogenies inferred from the majority of these genes are consistent with a chlorophyte green algal origin. However, a significant number of genes ( approximately 21%) show signs of having been acquired by lateral gene transfer from numerous other sources: streptophyte algae, red algae (or algae with red algal endosymbionts), as well as bacteria. The chlorarachniophyte plastid proteome may therefore be regarded as a mosaic derived from various organisms in addition to the ancestral chlorophyte plastid. In contrast, the homologous genes from the chlorophyte Chlamydomonas reinhardtii do not show any indications of lateral gene transfer. This difference is likely a reflection of the mixotrophic nature of Bigelowiella (i.e., it is photosynthetic and phagotrophic), whereas Chlamydomonas is strictly autotrophic. These results underscore the importance of lateral gene transfer in contributing foreign proteins to eukaryotic cells and their organelles, and also suggest that its impact can vary from lineage to lineage.
Figures



Comment in
-
Horizontal gene transfer in eukaryotic algal evolution.Proc Natl Acad Sci U S A. 2003 Jun 24;100(13):7419-20. doi: 10.1073/pnas.1533212100. Epub 2003 Jun 16. Proc Natl Acad Sci U S A. 2003. PMID: 12810941 Free PMC article. No abstract available.
References
-
- Delwiche, C. F. & Palmer, J. D. (1997) in Origins of Algae and Their Plastids, ed. Bhattacharya, D. (Springer, New York), pp. 53-86.
-
- McFadden, G. I. (2001) J. Phycol. 37, 951-959.
-
- Moreira, D., Le Guyader, H. & Phillippe, H. (2000) Nature 405, 69-72. - PubMed
-
- Archibald, J. M. & Keeling, P. J. (2002) Trends Genet. 18, 577-584. - PubMed
-
- Hibberd, D. J. & Norris, R. E. (1984) J. Phycol. 20, 310-330.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
