

Release of endogenous anti-inflammatory complement regulators FHL-1 and factor H protects synovial fibroblasts during rheumatoid arthritis
- PMID: 12780697
- PMCID: PMC1808733
- DOI: 10.1046/j.1365-2249.2003.02173.x
Release of endogenous anti-inflammatory complement regulators FHL-1 and factor H protects synovial fibroblasts during rheumatoid arthritis
Abstract
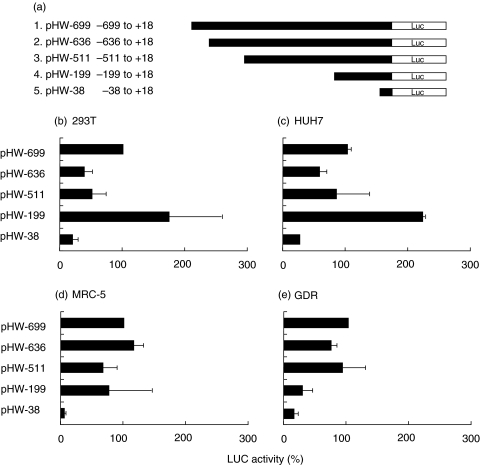
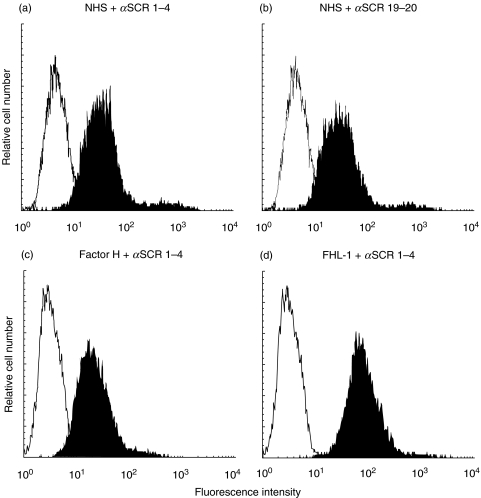
Rheumatoid arthritis is a chronic inflammatory disease of unknown aetiology predominantly affecting cells and tissues of synovial joints. Here we show that the two important complement regulators FHL-1 and factor H play a protective anti-inflammatory role in rheumatoid arthritis. Expression analyses at the mRNA- and protein level show in vitro expression and secretion of both regulators by synovial fibroblasts derived from patients with rheumatoid arthritis. Similarly the two regulators are synthesized in vivo in diseased synovial tissue, and in particular synovial lining cells express high levels of FHL-1. The anti-inflammatory role of these regulators in rheumatoid arthritis is highlighted by their induction with IFN-gamma and dexamethasone, whilst the pro-inflammatory cytokine TNF-alpha had no effect. Transient transfection experiments with various FHL-1/factor H promoter-luciferase reporter constructs into cells of distinct origin show independent cell and tissue specific promoter regulated transcription of these two regulators. The inducible expression, specifically of FHL-1 has physiological consequences. By binding directly to surfaces the released proteins protect cells from inflammatory damage and complement-mediated cell lysis. This study shows a novel protective and anti-inflammatory role of the two important complement regulators FHL-1 and factor H in rheumatoid arthritis and suggests a disease controlling role of the two proteins.
Figures






References
-
- Müller-Eberhard HJ, Schreiber RD. Molecular biology and chemistry of the alternative pathway of complement. Adv Immunol. 1980;29:1–53. - PubMed
-
- Walport MJ. Complement. N Engl J Med. 2001;344:1140–4. - PubMed
-
- Rus HG, Niculescu FI, Shin ML. Role of the C5b-9 complement complex in cell cycle and apoptosis. Immunol Rev. 2001;180:49–55. - PubMed
-
- Zipfel PF, Skerka C. FHL-1: a human complement and immune regulator with cell-adhesive function. Immunol Today. 1999;20:135–40. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
