

TRF1 is degraded by ubiquitin-mediated proteolysis after release from telomeres
- PMID: 12782650
- PMCID: PMC196064
- DOI: 10.1101/gad.1077103
TRF1 is degraded by ubiquitin-mediated proteolysis after release from telomeres
Abstract
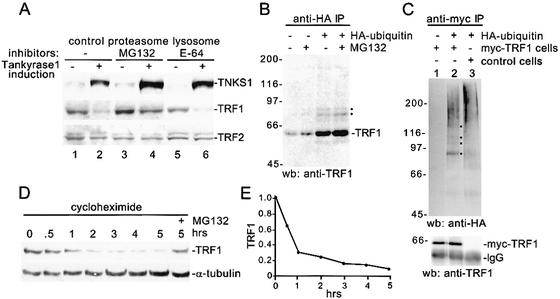
Mammalian telomeres are coated by the sequence-specific, DNA-binding protein, TRF1, a negative regulator of telomere length. Previous results showed that ADP-ribosylation of TRF1 by tankyrase 1 released TRF1 from telomeres and promoted telomere elongation. We now show that loss of TRF1 from telomeres results in ubiquitination and degradation of TRF1 by the proteasome and that degradation is required to keep TRF1 off telomeres. Ubiquitination of TRF1 is regulated by its telomere-binding status; only the telomere-unbound form of TRF1 is ubiquitinated. Our findings suggest a novel mechanism of sequential post translational modification of TRF1 (ADP-ribosylation and ubiquitination) for regulating access of telomerase to telomeres.
Figures




References
-
- Ancelin K., Brunori, M., Bauwens, S., Koering, C.E., Brun, C., Ricoul, M., Pommier, J.P., Sabatier, L., and Gilson, E. 2002. Targeting assay to study the cis functions of human telomeric proteins: Evidence for inhibition of telomerase by TRF1 and for activation of telomere degradation by TRF2. Mol. Cell. Biol. 22: 3474–3487. - PMC - PubMed
-
- Bilaud T., Brun, C., Ancelin, K., Koering, C.E., Laroche, T., and Gilson, E. 1997. Telomeric localization of TRF2, a novel human telobox protein. Nat. Genet. 17: 236–239. - PubMed
-
- Broccoli D., Smogorzewska, A., Chong, L., and de Lange, T. 1997. Human telomeres contain two distinct Myb-related proteins, TRF1 and TRF2. Nat. Genet. 17: 231–235. - PubMed
-
- Cenci G., Rawson, R.B., Belloni, G., Castrillon, D.H., Tudor, M., Petrucci, R., Goldberg, M.L., Wasserman, S.A., and Gatti, M. 1997. UbcD1, a Drosophila ubiquitin-conjugating enzyme required for proper telomere behavior. Genes & Dev. 11: 863–875. - PubMed
-
- Chong L., van Steensel, B., Broccoli, D., Erdjument-Bromage, H., Hanish, J., Tempst, P., and de Lange, T. 1995. A human telomeric protein. Science 270: 1663–1667. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases