

MIF signal transduction initiated by binding to CD74
- PMID: 12782713
- PMCID: PMC2193907
- DOI: 10.1084/jem.20030286
MIF signal transduction initiated by binding to CD74
Abstract
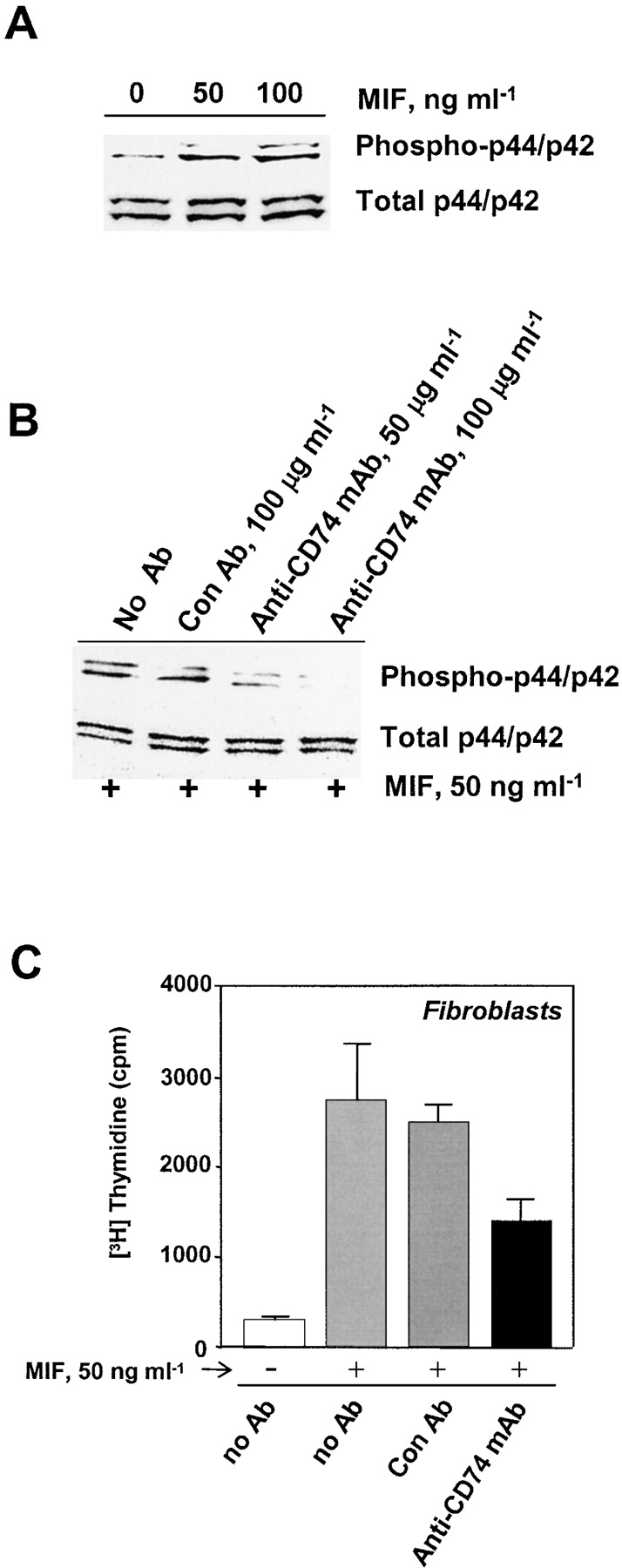
Macrophage migration inhibitory factor (MIF) accounts for one of the first cytokine activities to have been described, and it has emerged recently to be an important regulator of innate and adaptive immunity. MIF is an upstream activator of monocytes/macrophages, and it is centrally involved in the pathogenesis of septic shock, arthritis, and other inflammatory conditions. The protein is encoded by a unique but highly conserved gene, and X-ray crystallography studies have shown MIF to define a new protein fold and structural superfamily. Although recent work has begun to illuminate the signal transduction pathways activated by MIF, the nature of its membrane receptor has not been known. Using expression cloning and functional analysis, we report herein that CD74, a Type II transmembrane protein, is a high-affinity binding protein for MIF. MIF binds to the extracellular domain of CD74, and CD74 is required for MIF-induced activation of the extracellular signal-regulated kinase-1/2 MAP kinase cascade, cell proliferation, and PGE2 production. A recombinant, soluble form of CD74 binds MIF with a dissociation constant of approximately 9 x 10-9 Kd, as defined by surface plasmon resonance (BIAcore analysis), and soluble CD74 inhibits MIF-mediated extracellular signal-regulated kinase activation in defined cell systems. These data provide a molecular basis for MIF's interaction with target cells and identify it as a natural ligand for CD74, which has been implicated previously in signaling and accessory functions for immune cell activation.
Figures


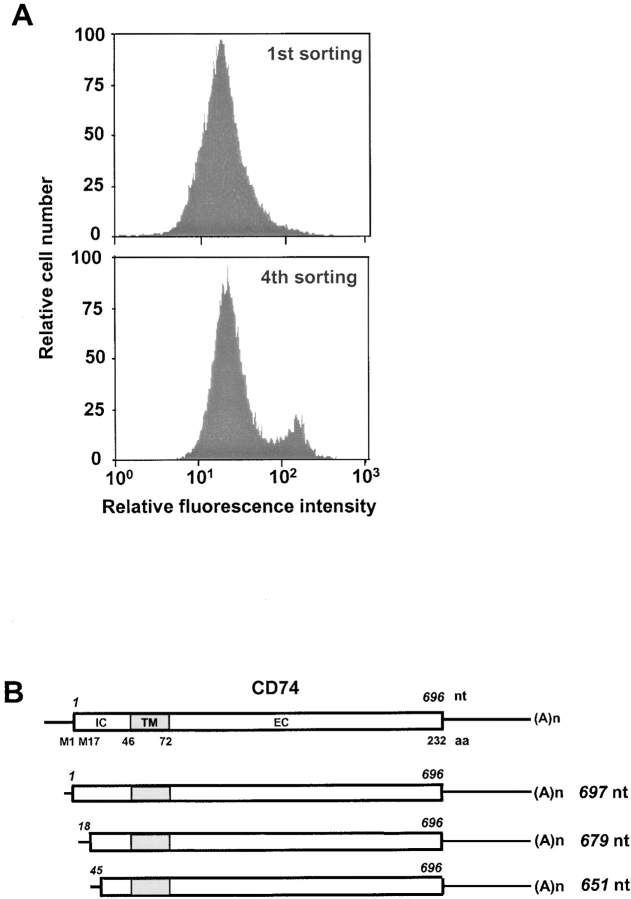

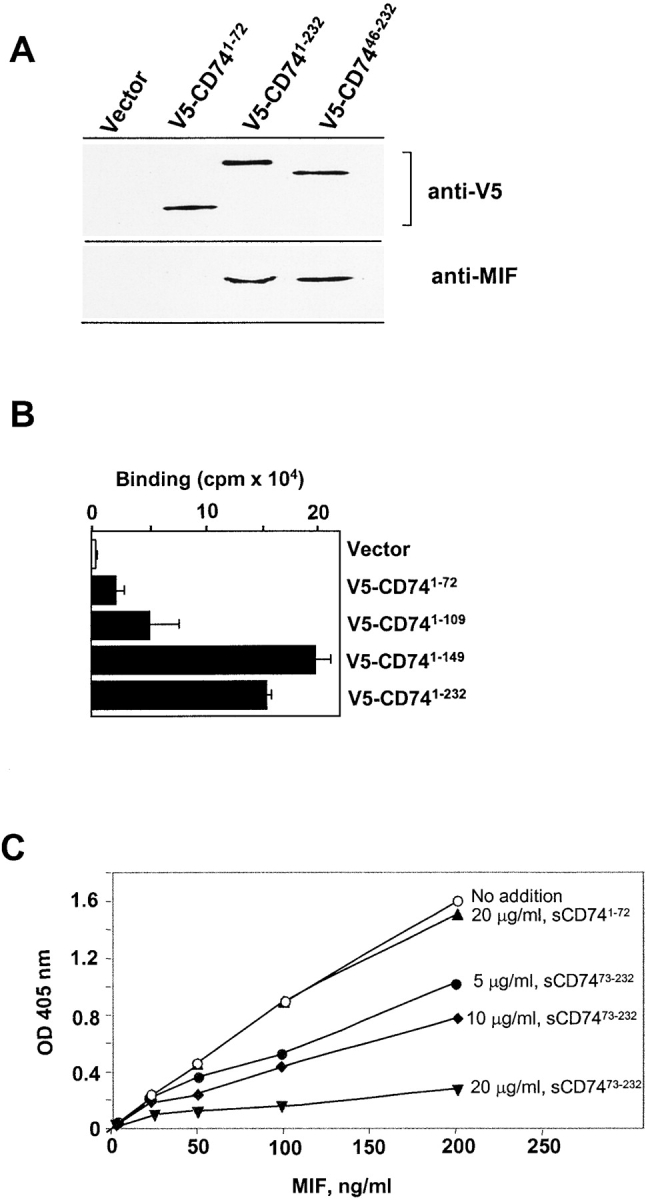
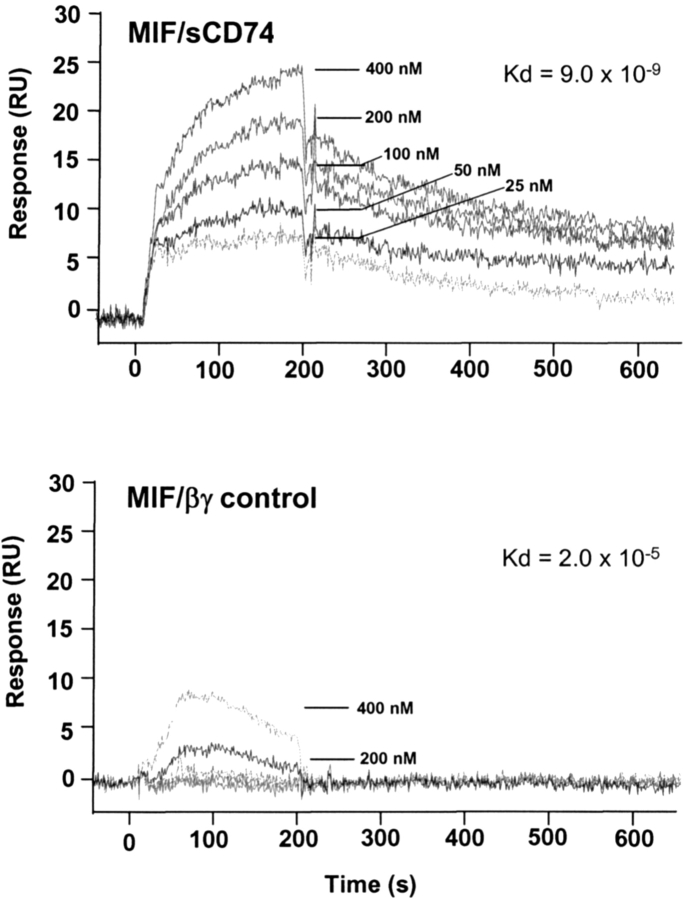

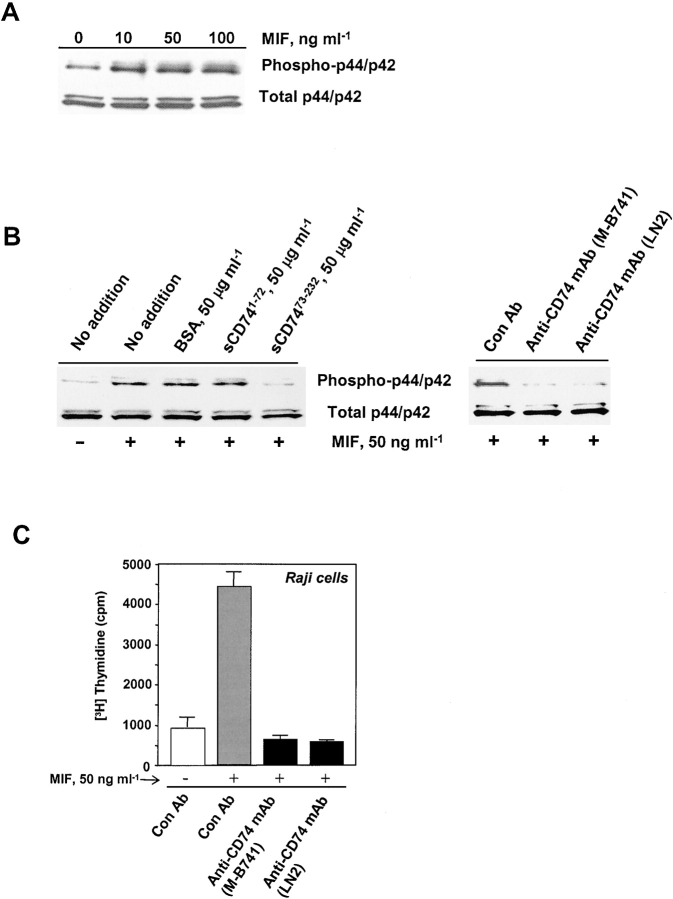
References
-
- George, M., and J.H. Vaughn. 1962. In vitro cell migration as a model for delayed hypersensitivity. Proc. Soc. Exp. Biol. Med. 111:514–521. - PubMed
-
- Bloom, B.R., and B. Bennett. 1966. Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science. 153:80–82. - PubMed
-
- Bernhagen, J., T. Calandra, R.A. Mitchell, S.B. Martin, K.J. Tracey, W. Voelter, K.R. Manogue, A. Cerami, and R. Bucala. 1993. MIF is a pituitary-derived cytokine that potentiates lethal endotoxaemia. Nature. 365:756–759. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
