

Protein kinase C theta affects Ca2+ mobilization and NFAT cell activation in primary mouse T cells
- PMID: 12782715
- PMCID: PMC2193906
- DOI: 10.1084/jem.20020234
Protein kinase C theta affects Ca2+ mobilization and NFAT cell activation in primary mouse T cells
Erratum in
- J Exp Med. 2003 Jul 7;198(1):183
Abstract
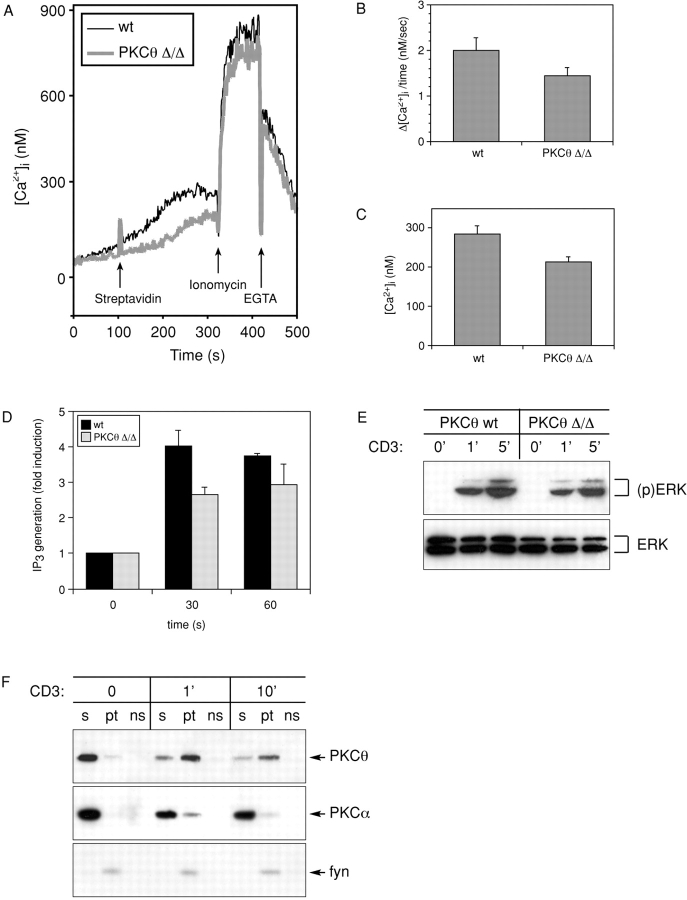
Protein kinase C (PKC)theta is an established component of the immunological synapse and has been implicated in the control of AP-1 and NF-kappaB. To study the physiological function of PKCtheta, we used gene targeting to generate a PKCtheta null allele in mice. Consistently, interleukin 2 production and T cell proliferative responses were strongly reduced in PKCtheta-deficient T cells. Surprisingly, however, we demonstrate that after CD3/CD28 engagement, deficiency of PKCtheta primarily abrogates NFAT transactivation. In contrast, NF-kappaB activation was only partially reduced. This NFAT transactivation defect appears to be secondary to reduced inositol 1,4,5-trisphosphate generation and intracellular Ca2+ mobilization. Our finding suggests that PKCtheta plays a critical and nonredundant role in T cell receptor-induced NFAT activation.
Figures






References
-
- Baier, G. 2003. The PKC gene module: molecular biosystematics to resolve its T cell functions. Immunol. Rev. 192:64–79. - PubMed
-
- Baier, G., D. Telford, L. Giampa, K.M. Coggeshall, G. Baier-Bitterlich, N. Isakov, and A. Altman. 1993. Molecular cloning and characterization of PKCθ, a novel member of the protein kinase C (PKC) gene family expressed predominantly in hematopoietic cells. J. Biol. Chem. 268:4997–5004. - PubMed
-
- Baier, G., G. Baier-Bitterlich, N. Meller, K.M. Coggeshall, L. Giampa, D. Telford, N. Isakov, and A. Altman. 1994. Expression and biochemical characterization of human PKCθ. Eur. J. Biochem. 225:195–203. - PubMed
-
- Wilda, M., N. Ghaffari-Tabrizi, I. Reisert, G. Utermann, G. Baier, and H. Hameister. 2001. Protein kinase C isoenzyme: selective expression pattern of PKCθ; during mouse development. Mech. Dev. 103:197–200. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
