

The Escherichia coli amidase AmiC is a periplasmic septal ring component exported via the twin-arginine transport pathway
- PMID: 12787347
- PMCID: PMC4428285
- DOI: 10.1046/j.1365-2958.2003.03511.x
The Escherichia coli amidase AmiC is a periplasmic septal ring component exported via the twin-arginine transport pathway
Abstract
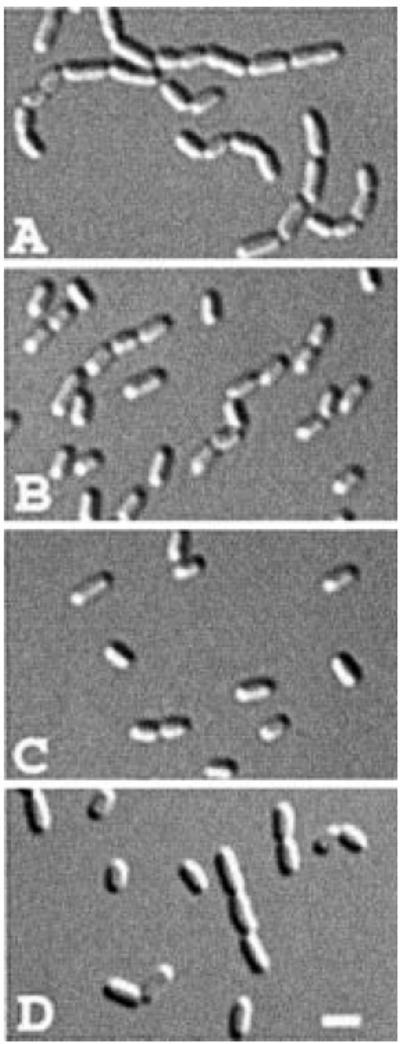
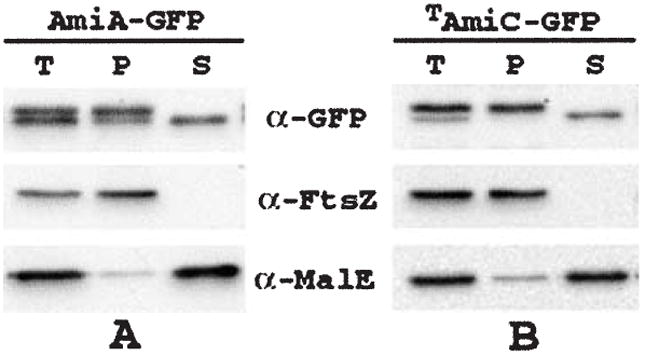
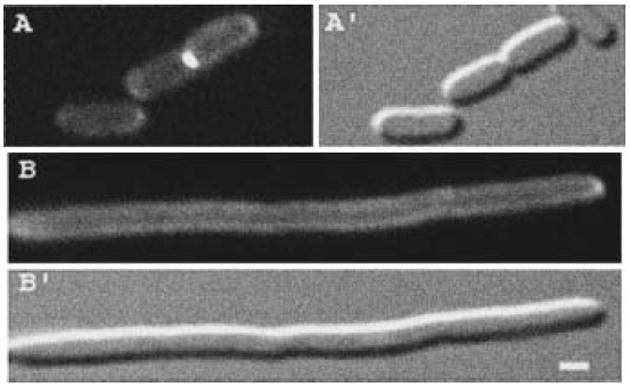
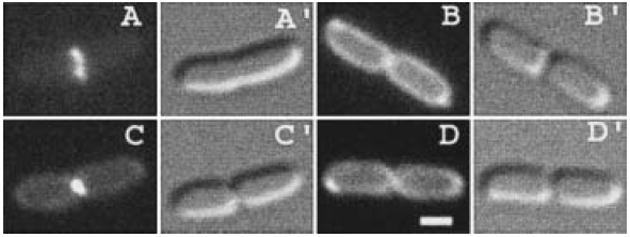
The N-acetylmuramoyl-l-alanine amidases of Escherichia coli (AmiA, B and C) are periplasmic enzymes that remove murein cross-links by cleaving the peptide moiety from N-acetylmuramic acid. Ami- cells form chains, indicating that the amidases help to split the septal murein. Interestingly, cells defective in the twin-arginine protein transport (Tat) pathway show a similar division defect. We find that both AmiA and AmiC are routed to the periplasm via Tat, providing an explanation for the Tat- division phenotype. Taking advantage of the ability of Tat to export prefolded (fluorescent) green fluorescent protein (GFP) to the periplasm, we sublocalized AmiA and AmiC in live cells using functional fusions to GFP. Interestingly, the periplasmic localization of the fusions differed markedly. AmiA-GFP appeared to be dispersed throughout the periplasm in all cells. AmiC-GFP similarly appeared throughout the periplasm in small cells, but was concentrated almost exclusively at the septal ring in constricting cells. Recruitment of AmiC to the ring was mediated by an N-terminal non-amidase targeting domain and required the septal ring component FtsN. AmiC therefore replaces FtsN as the latest known recruit to the septal ring and is the first entirely periplasmic component to be localized.
Figures




References
-
- Adam M, Fraipont C, Rhazi N, Nguyen-Disteche M, Lakaye B, Frere JM, et al. The bimodular G57–V577 polypeptide chain of the class B penicillin-binding protein 3 of Escherichia coli catalyzes peptide bond formation from thiolesters and does not catalyze glycan chain polymerization from the lipid II intermediate. J Bacteriol. 1997;179:6005–6009. - PMC - PubMed
-
- Addinall SG, Cao C, Lutkenhaus J. FtsN, a late recruit to the septum in Escherichia coli. Mol Microbiol. 1997;25:303–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
