

The intestinal protozoan parasite Entamoeba histolytica contains 20 cysteine protease genes, of which only a small subset is expressed during in vitro cultivation
- PMID: 12796295
- PMCID: PMC161451
- DOI: 10.1128/EC.2.3.501-509.2003
The intestinal protozoan parasite Entamoeba histolytica contains 20 cysteine protease genes, of which only a small subset is expressed during in vitro cultivation
Abstract
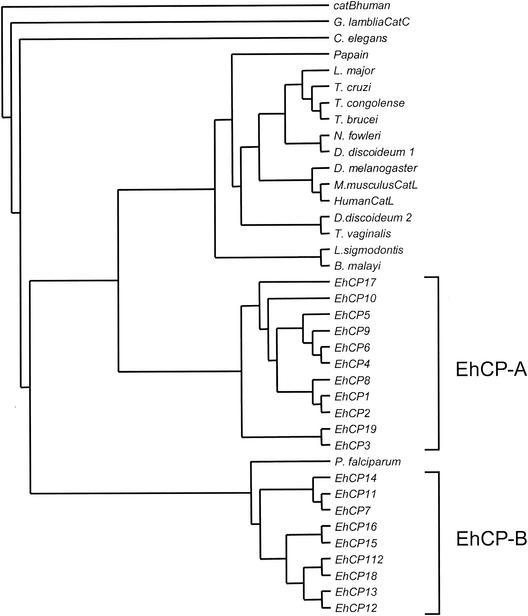
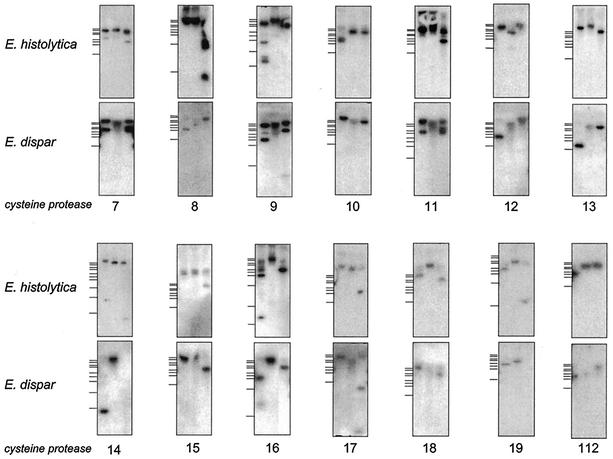
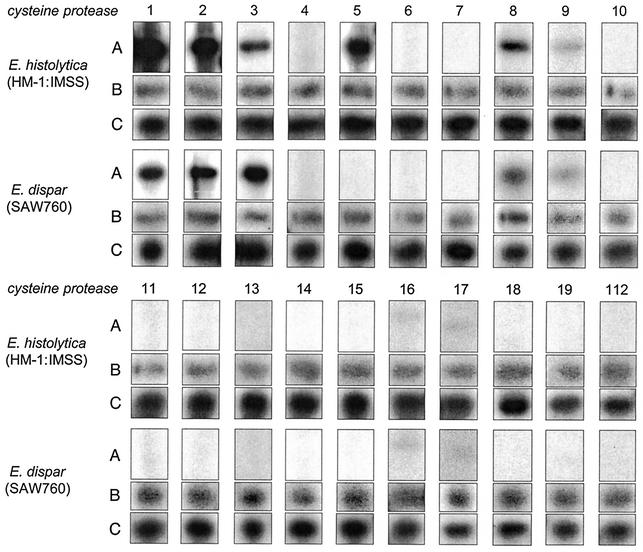
Cysteine proteases are known to be important pathogenicity factors of the protozoan parasite Entamoeba histolytica. So far, a total of eight genes coding for cysteine proteases have been identified in E. histolytica, two of which are absent in the closely related nonpathogenic species E. dispar. However, present knowledge is restricted to enzymes expressed during in vitro cultivation of the parasite, which might represent only a subset of the entire repertoire. Taking advantage of the current E. histolytica genome-sequencing efforts, we analyzed databases containing more than 99% of all ameba gene sequences for the presence of cysteine protease genes. A total of 20 full-length genes was identified (including all eight genes previously reported), which show 10 to 86% sequence identity. The various genes obviously originated from two separate ancestors since they form two distinct clades. Despite cathepsin B-like substrate specificities, all of the ameba polypeptides are structurally related to cathepsin L-like enzymes. None of the previously described enzymes but 7 of the 12 newly identified proteins are unique compared to cathepsins of higher eukaryotes in that they are predicted to have transmembrane or glycosylphosphatidylinositol anchor attachment domains. Southern blot analysis revealed that orthologous sequences for all of the newly identified proteases are present in E. dispar. Interestingly, the majority of the various cysteine protease genes are not expressed in E. histolytica or E. dispar trophozoites during in vitro cultivation. Therefore, it is likely that at least some of these enzymes are required for infection of the human host and/or for completion of the parasite life cycle.
Figures




References
-
- Ankri, S., T. Stolarsky, and D. Mirelman. 1998. Antisense inhibition of expression of cysteine proteinases does not affect Entamoeba histolytica cytopathic or haemolytic activity but inhibits phagocytosis. Mol. Microbiol. 28:777-785. - PubMed
-
- Avila, E. E., and J. Calderon. 1993. Entamoeba histolytica trophozoites: a surface-associated cysteine protease. Exp. Parasitol. 76:232-241. - PubMed
-
- Barrett, A. J. 1998. Cysteine peptidase. In A. J. Barrett, N. D. Rawlings, and J. F. Woessner (ed.), Handbook of proteolytic enzymes. Academic Press, Inc., San Diego, Calif.
-
- Bassi, D. E., H. Mahloogi, and A. J. Klein-Szanto. 2000. The proprotein convertases furin and PACE4 play a significant role in tumor progression. Mol. Carcinog. 28:63-69. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
