

RNA polymerase I transcribes procyclin genes and variant surface glycoprotein gene expression sites in Trypanosoma brucei
- PMID: 12796299
- PMCID: PMC161450
- DOI: 10.1128/EC.2.3.542-551.2003
RNA polymerase I transcribes procyclin genes and variant surface glycoprotein gene expression sites in Trypanosoma brucei
Abstract
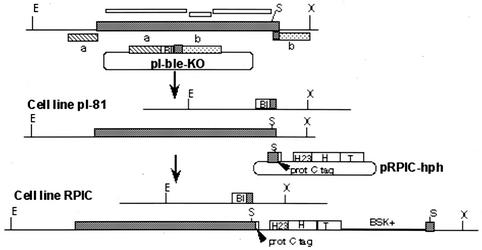
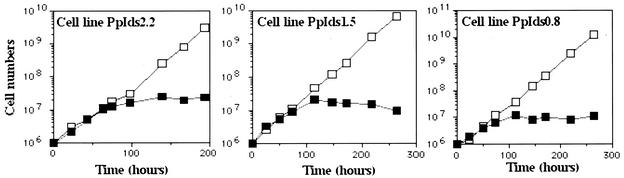
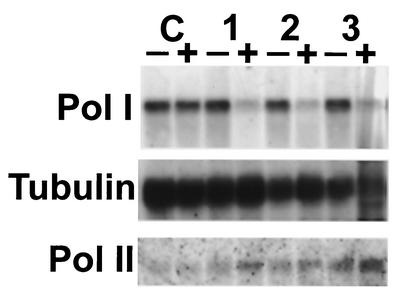
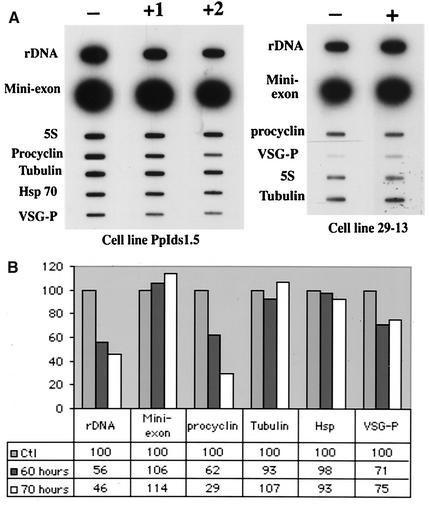
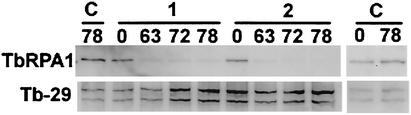
In eukaryotes, RNA polymerase (pol) I exclusively transcribes the large rRNA gene unit (rDNA) and mRNA is synthesized by RNA pol II. The African trypanosome, Trypanosoma brucei, represents an exception to this rule. In this organism, transcription of genes encoding the variant surface glycoprotein (VSG) and the procyclins is resistant to alpha-amanitin, indicating that it is mediated by RNA pol I, while other protein-coding genes are transcribed by RNA pol II. To obtain firm proof for this concept, we generated a T. brucei cell line which exclusively expresses protein C epitope-tagged RNA pol I. Using an anti-protein C immunoaffinity matrix, we specifically depleted RNA pol I from transcriptionally active cell extracts. The depletion of RNA pol I impaired in vitro transcription initiated at the rDNA promoter, the GPEET procyclin gene promoter, and a VSG gene expression site promoter but did not affect transcription from the spliced leader (SL) RNA gene promoter. Fittingly, induction of RNA interference against the RNA pol I largest subunit in insect-form trypanosomes significantly reduced the relative transcriptional efficiency of rDNA, procyclin genes, and VSG expression sites in vivo whereas that of SL RNA, alphabeta-tubulin, and heat shock protein 70 genes was not affected. Our studies unequivocally show that T. brucei harbors a multifunctional RNA pol I which, in addition to transcribing rDNA, transcribes procyclin genes and VSG gene expression sites.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
