

A novel mechanism of myocyte degeneration involving the Ca2+-permeable growth factor-regulated channel
- PMID: 12796481
- PMCID: PMC2172975
- DOI: 10.1083/jcb.200301101
A novel mechanism of myocyte degeneration involving the Ca2+-permeable growth factor-regulated channel
Abstract
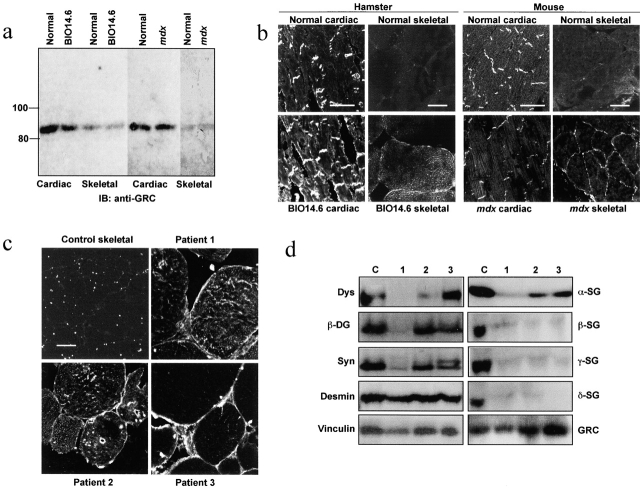
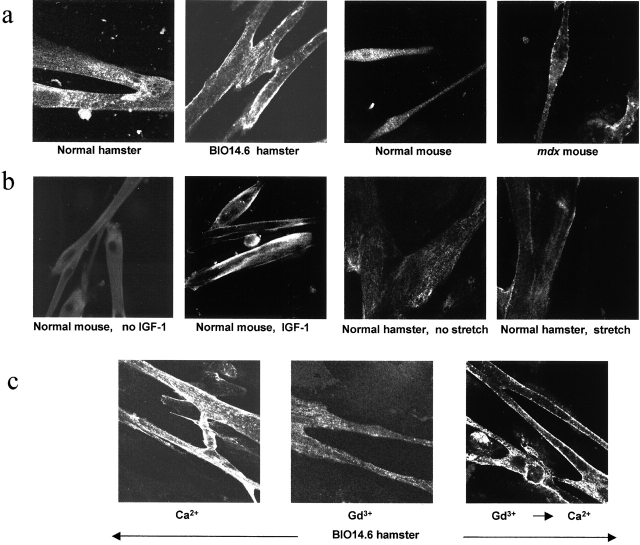
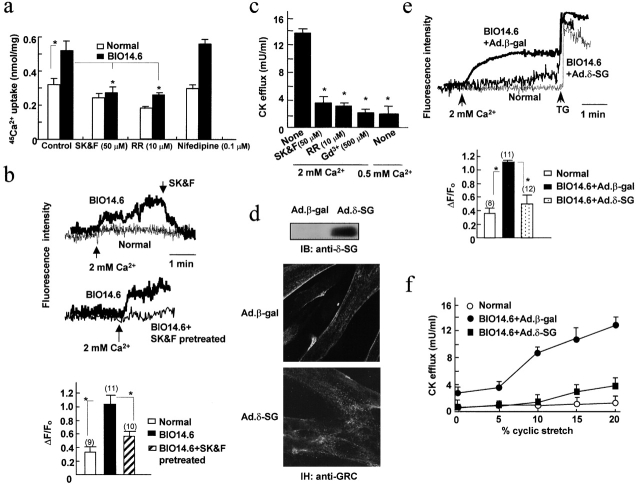
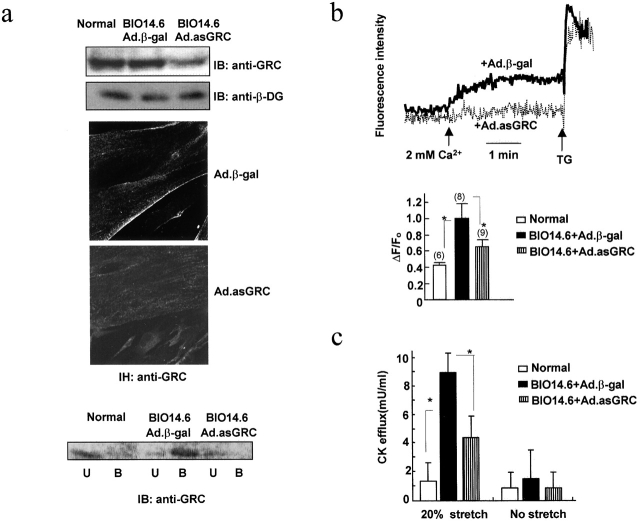
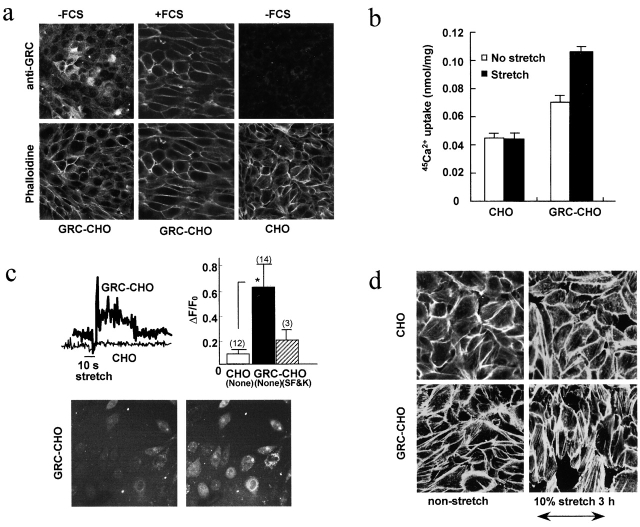
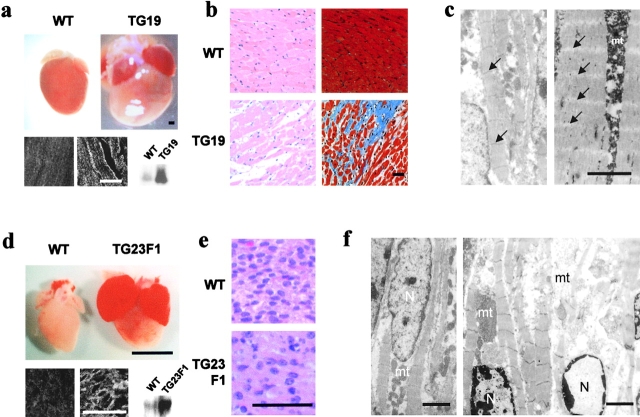
Disruption of the dystrophin-glycoprotein complex caused by genetic defects of dystrophin or sarcoglycans results in muscular dystrophy and/or cardiomyopathy in humans and animal models. However, the key early molecular events leading to myocyte degeneration remain elusive. Here, we observed that the growth factor-regulated channel (GRC), which belongs to the transient receptor potential channel family, is elevated in the sarcolemma of skeletal and/or cardiac muscle in dystrophic human patients and animal models deficient in dystrophin or delta-sarcoglycan. However, total cell GRC does not differ markedly between normal and dystrophic muscles. Analysis of the properties of myotubes prepared from delta-sarcoglycan-deficient BIO14.6 hamsters revealed that GRC is activated in response to myocyte stretch and is responsible for enhanced Ca2+ influx and resultant cell damage as measured by creatine phosphokinase efflux. We found that cell stretch increases GRC translocation to the sarcolemma, which requires entry of external Ca2+. Consistent with these findings, cardiac-specific expression of GRC in a transgenic mouse model produced cardiomyopathy due to Ca2+ overloading, with disease expression roughly parallel to sarcolemmal GRC levels. The results suggest that GRC is a key player in the pathogenesis of myocyte degeneration caused by dystrophin-glycoprotein complex disruption.
Figures
References
-
- Alderton, J.M., and R.A. Steinhardt. 2000. a. Calcium influx through calcium leak channels is responsible for the elevated levels of calcium-dependent proteolysis in dystrophic myotubes. J. Biol. Chem. 275:9452–9460. - PubMed
-
- Alderton, J.M., and R.A. Steinhardt. 2000. b. How calcium influx through calcium leak channels is responsible for the elevated levels of calcium-dependent proteolysis in dystrophic myotubes. Trends Cardiovasc Med. 10:268–272. - PubMed
-
- Bajusz, E., F. Homburger, J.R. Baker, and P. Bogdonoff. 1969. Dissociation of factors influencing myocardial degeneration and generalized cardiocirculatory failure. Ann. NY Acad. Sci. 156:396–420. - PubMed
-
- Bertrand, B., S. Wakabayashi, S. Ikeda, J. Pouysségur, and M. Shigekawa. 1994. The Na+/H+ exchanger isoform 1 (NHE1) is a novel member of the calmodulin-binding proteins. Identification and characterization of calmodulin-binding sites. J. Biol. Chem. 269:13703–13709. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
