

Neurexin mediates the assembly of presynaptic terminals
- PMID: 12796785
- PMCID: PMC1646425
- DOI: 10.1038/nn1074
Neurexin mediates the assembly of presynaptic terminals
Abstract
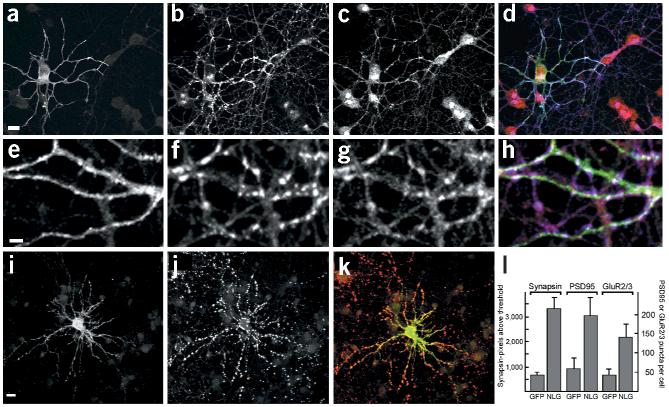
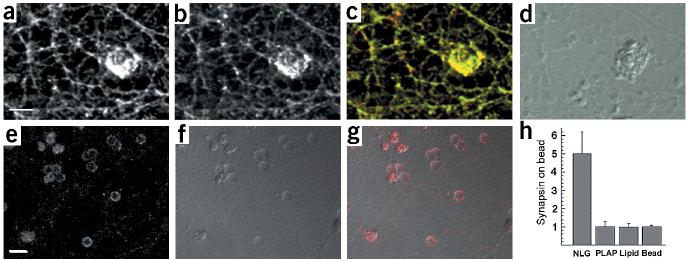
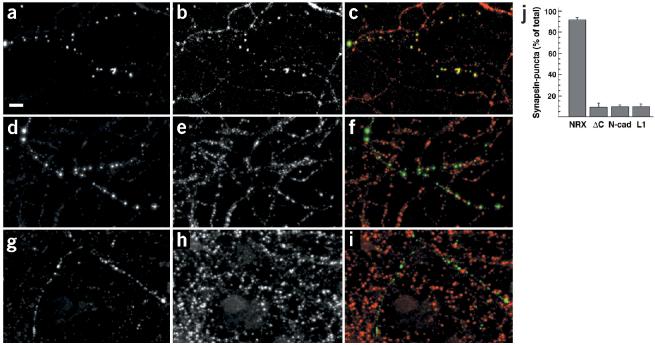
Neurexins are a large family of proteins that act as neuronal cell-surface receptors. The function and localization of the various neurexins, however, have not yet been clarified. Beta-neurexins are candidate receptors for neuroligin-1, a postsynaptic membrane protein that can trigger synapse formation at axon contacts. Here we report that neurexins are concentrated at synapses and that purified neuroligin is sufficient to cluster neurexin and to induce presynaptic differentiation. Oligomerization of neuroligin is required for its function, and we find that beta-neurexin clustering is sufficient to trigger the recruitment of synaptic vesicles through interactions that require the cytoplasmic domain of neurexin. We propose a two-step model in which postsynaptic neuroligin multimers initially cluster axonal neurexins. In response to this clustering, neurexins nucleate the assembly of a cytoplasmic scaffold to which the exocytotic apparatus is recruited.
Figures
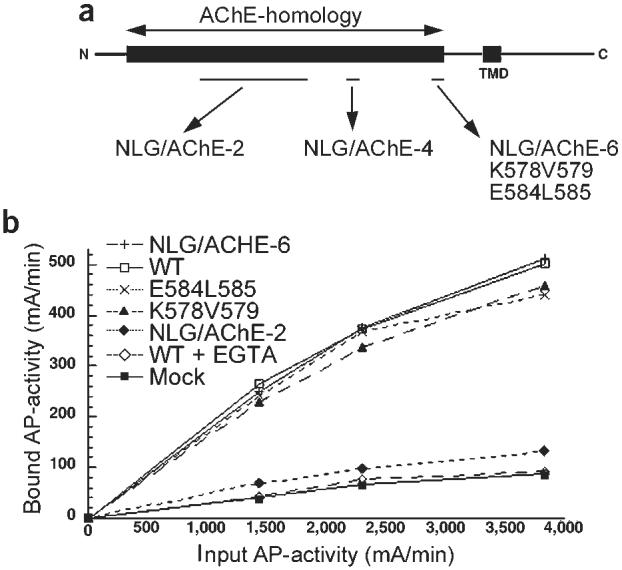
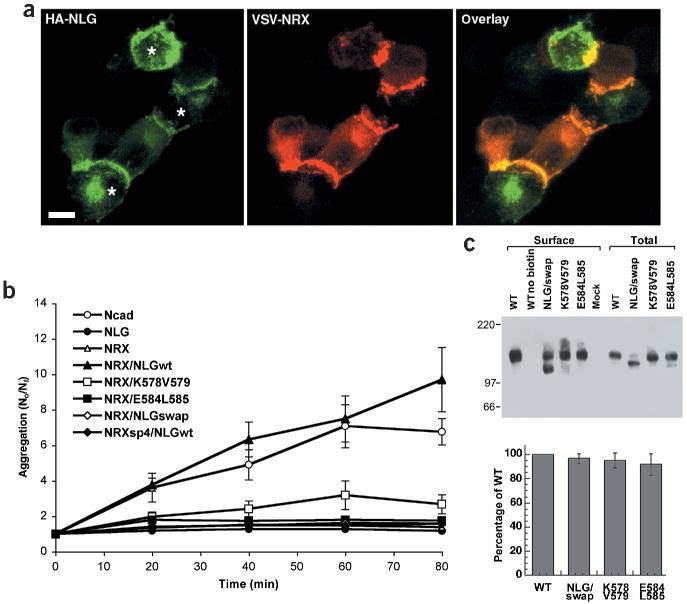
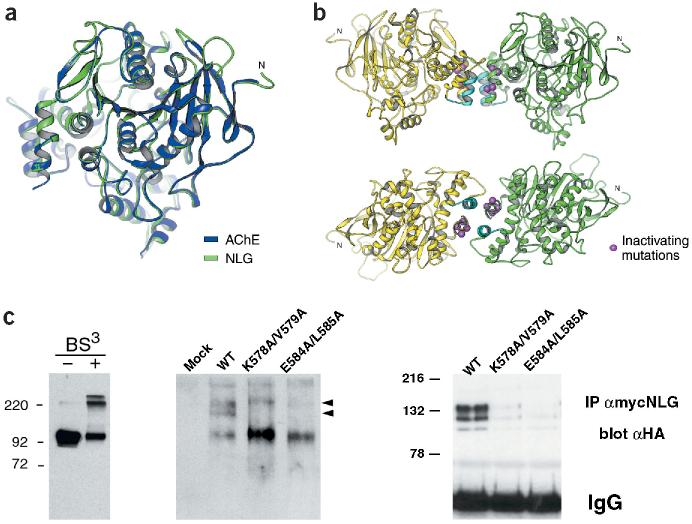
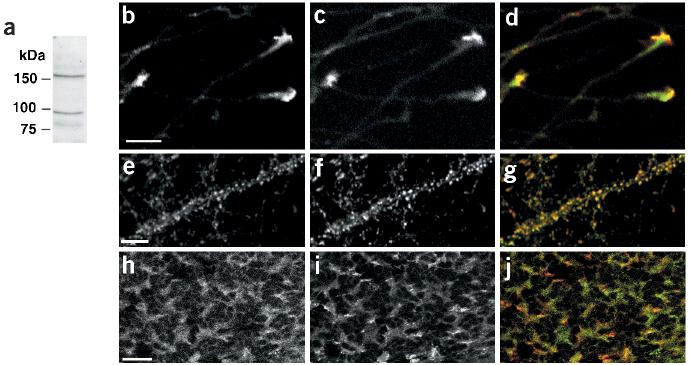
References
-
- Sanes JR, Lichtman JW. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 1999;22:389–442. - PubMed
-
- Garner CC, Zhai RG, Gundelfinger ED, Ziv NE. Molecular mechanisms of CNS synaptogenesis. Trends Neurosci. 2002;25:243–251. - PubMed
-
- Scheiffele P. Cell-cell signaling during synapse formation in the CNS. Annu. Rev. Neurosci. 2003;26:485–508. - PubMed
-
- Ichtchenko K, et al. Neuroligin 1: a splice site-specific ligand for beta-neurexins. Cell. 1995;81:435–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
