

Interaction of the bullous pemphigoid antigen 1 (BP230) and desmoplakin with intermediate filaments is mediated by distinct sequences within their COOH terminus
- PMID: 12802069
- PMCID: PMC165091
- DOI: 10.1091/mbc.e02-08-0548
Interaction of the bullous pemphigoid antigen 1 (BP230) and desmoplakin with intermediate filaments is mediated by distinct sequences within their COOH terminus
Abstract
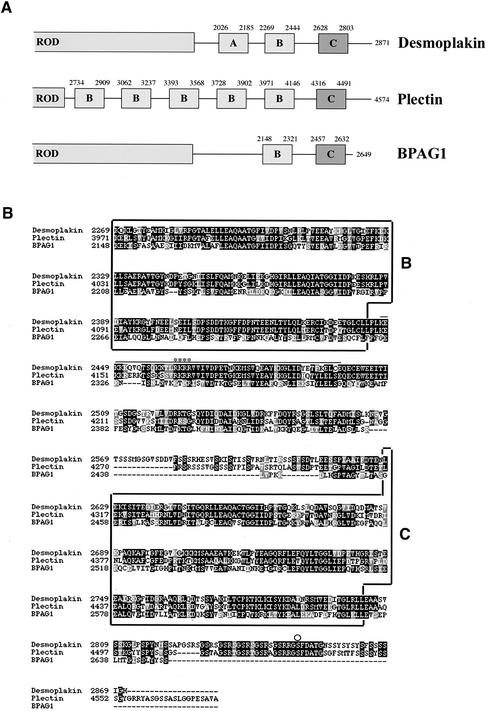
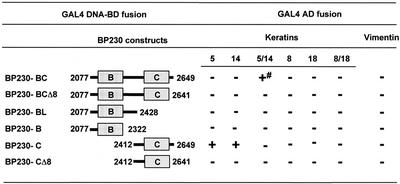
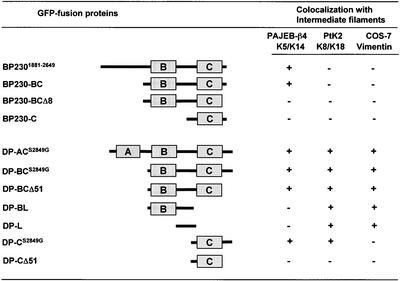
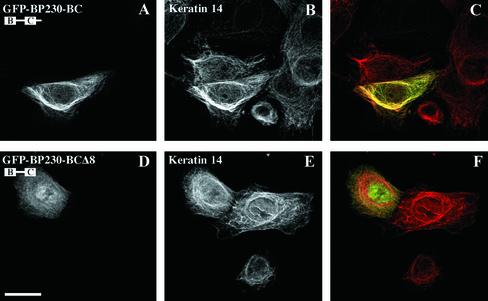
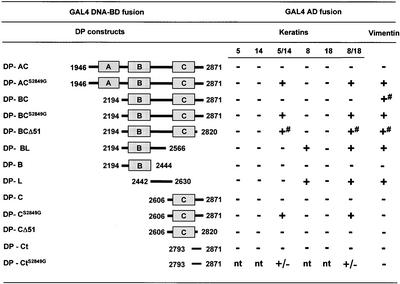
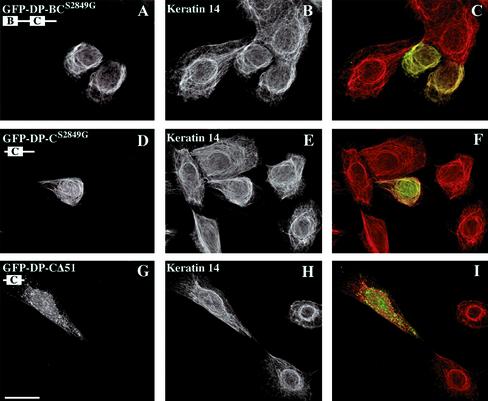
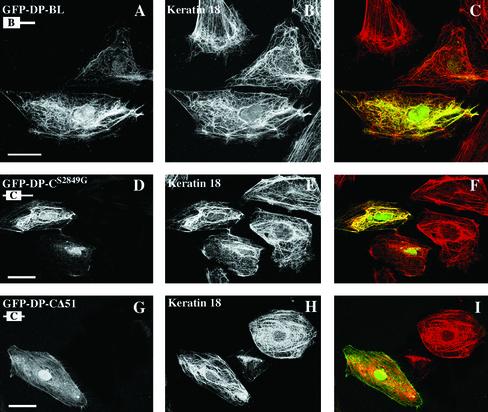
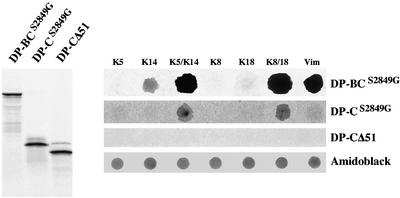
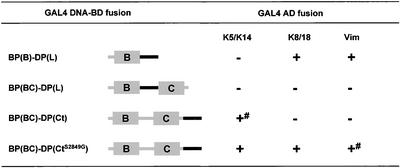
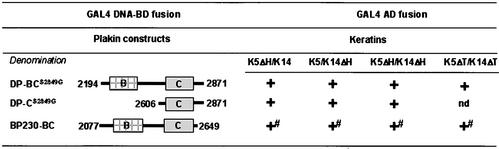
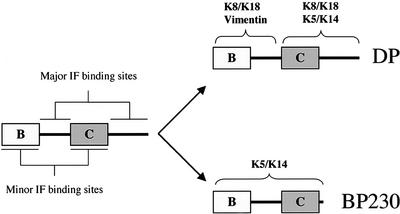
The bullous pemphigoid antigen 1 (BP230) and desmoplakin (DP) are members of the plakin protein family of cytolinkers. Despite their homology, their COOH termini selectively bind distinct intermediate filaments (IFs). We studied sequences within their COOH termini required for their interaction with the epidermal keratins K5/K14, the simple epithelial keratins K8/K18, and type III IF vimentin by yeast three-hybrid, cell transfection, and overlay assays. The results indicate that BP230 interacts with K5/K14 but not with K8/K18 or vimentin via a region encompassing both the B and C subdomains and the COOH extremity, including a COOH-terminal eight-amino-acid stretch. In contrast, the C subdomain with the COOH-terminal extremity of DP interacts with K5/K14 and K8/K18, and its linker region is able to associate with K8/K18 and vimentin. Furthermore, the potential of DP to interact with IF proteins in yeast seems to be regulated by phosphorylation of Ser 2849 within its COOH terminus. Strikingly, BP230 and DP interacted with cytokeratins only when both type I and type II keratins were present. The head and tail domains of K5/K14 keratins were dispensable for their interaction with BP230 or DP. On the basis of our findings, we postulate that (1) the binding specificity of plakins for various IF proteins depends on their linker region between the highly homologous B and C subdomains and their COOH extremity and (2) the association of DP and BP230 with both epidermal and simple keratins is critically affected by the tertiary structure induced by heterodimerization and involves recognition sites located primarily in the rod domain of these keratins.
Figures
References
-
- Angst, B.D., Nilles, L.A., and Green, K.J. (1990). Desmoplakin II expression is not restricted to stratified epithelia. J. Cell Sci. 97, 247–257. - PubMed
-
- Arnemann, J., Sullivan, K.H., Magee, A.I., King, I.A., and Buxton, R.S. (1993). Stratification-related expression of isoforms of the desmosomal cadherins in human epidermis. J. Cell Sci. 104, 741–750. - PubMed
-
- Borradori, L., and Sonnenberg, A. (1999). Structure and function of hemidesmosomes: more than simple adhesion complexes. J. Invest. Dermatol. 112, 411–418. - PubMed
-
- Brown, A., Bernier, G., Mathieu, M., Rossant, J., and Kothary, R. (1995). The mouse dystonia musculorum gene is a neural isoform of bullous pemphigoid antigen 1. Nat. Genet. 10, 301–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
