

LMP1, a viral relative of the TNF receptor family, signals principally from intracellular compartments
- PMID: 12805217
- PMCID: PMC162136
- DOI: 10.1093/emboj/cdg284
LMP1, a viral relative of the TNF receptor family, signals principally from intracellular compartments
Abstract
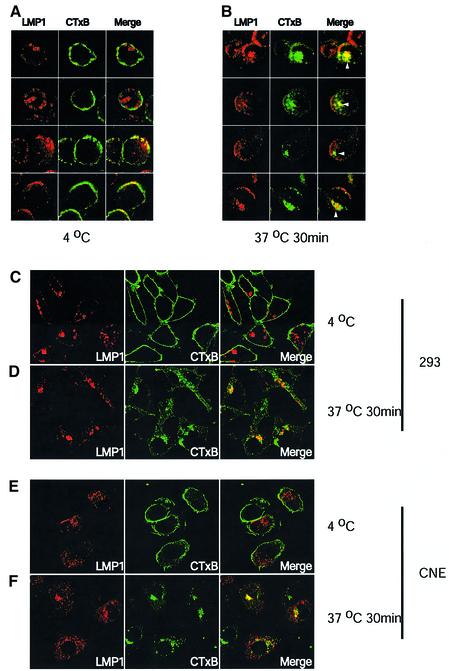
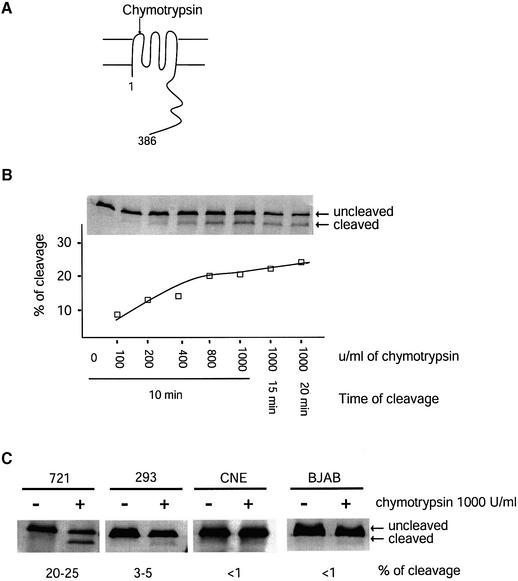
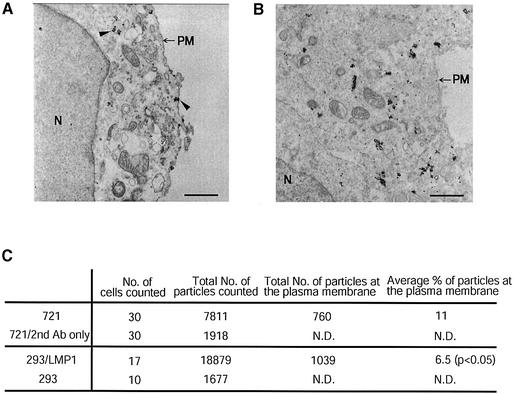
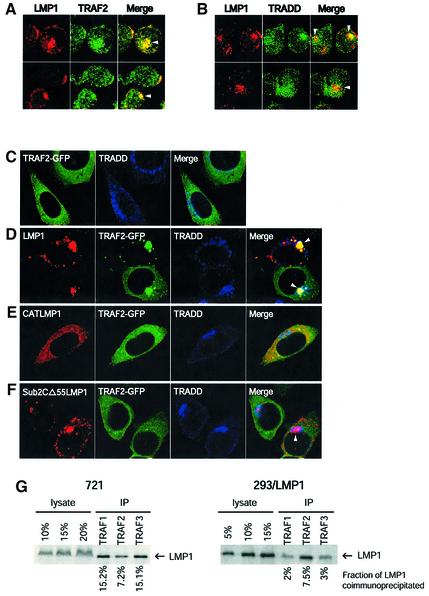
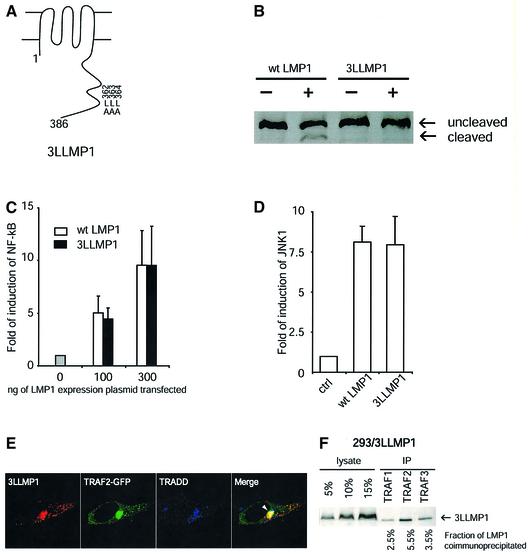
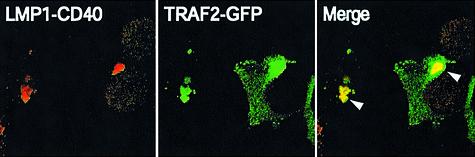
Latent membrane protein 1 (LMP1) is an Epstein-Barr virus (EBV)-encoded, ligand-independent receptor that mimics CD40. We report here that LMP1 signals principally from intracellular compartments. LMP1 associates simultaneously with lipid rafts and with its signaling molecules, tumor necrosis factor-receptor (TNF-R)-associated factors (TRAFs) and TNF-R1-associated death domain protein (TRADD) intracellularly, although it can be detected at low levels at the plasma membrane, indicating that most of LMP1's signaling complex resides in intracellular compartments. LMP1's signaling is independent of its accumulation at the plasma membrane in different cells, and as demonstrated by a mutant of LMP1 which has significantly reduced localization at the plasma membrane yet signals as efficiently as does wild-type LMP1. The fusion of the transmembrane domain of LMP1 to signaling domains of CD40, TNF-R1 and Fas activates their signaling; we demonstrate that a fusion of LMP1 with CD40 recruits TRAF2 intracellularly. Our results imply that members of the TNF-R family can signal from intracellular compartments containing lipid rafts and may do so when they act in autocrine loops.
Figures
Similar articles
-
Latent membrane protein 1 of Epstein-Barr virus mimics a constitutively active receptor molecule.EMBO J. 1997 Oct 15;16(20):6131-40. doi: 10.1093/emboj/16.20.6131. EMBO J. 1997. PMID: 9359753 Free PMC article.
-
A novel dominant-negative mutant form of Epstein-Barr virus latent membrane protein-1 (LMP1) selectively and differentially impairs LMP1 and TNF signaling pathways.Oncogene. 2004 Apr 8;23(15):2681-93. doi: 10.1038/sj.onc.1207432. Oncogene. 2004. PMID: 14767477
-
LMP1 signal transduction differs substantially from TNF receptor 1 signaling in the molecular functions of TRADD and TRAF2.EMBO J. 1999 May 4;18(9):2511-21. doi: 10.1093/emboj/18.9.2511. EMBO J. 1999. PMID: 10228165 Free PMC article.
-
Pursuing different 'TRADDes': TRADD signaling induced by TNF-receptor 1 and the Epstein-Barr virus oncoprotein LMP1.Biol Chem. 2008 Oct;389(10):1261-71. doi: 10.1515/BC.2008.144. Biol Chem. 2008. PMID: 18713013 Review.
-
[Signal transduction and biological characteristics of EB virus-encoded latent membrane protein 1 and its correlation with CD40].Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2004 Oct;26(5):585-90. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2004. PMID: 15562778 Review. Chinese.
Cited by
-
Exosomes derived from Epstein-Barr virus-infected cells are internalized via caveola-dependent endocytosis and promote phenotypic modulation in target cells.J Virol. 2013 Sep;87(18):10334-47. doi: 10.1128/JVI.01310-13. Epub 2013 Jul 17. J Virol. 2013. PMID: 23864627 Free PMC article.
-
Gene profiling of Epstein-Barr Virus and human endogenous retrovirus in peripheral blood mononuclear cells of SLE patients: immune response implications.Sci Rep. 2024 Aug 30;14(1):20236. doi: 10.1038/s41598-024-70913-6. Sci Rep. 2024. PMID: 39215087 Free PMC article.
-
Transmembrane Domains Mediate Intra- and Extracellular Trafficking of Epstein-Barr Virus Latent Membrane Protein 1.J Virol. 2018 Aug 16;92(17):e00280-18. doi: 10.1128/JVI.00280-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29950415 Free PMC article.
-
Epstein Barr Virus Associated Lymphomas and Epithelia Cancers in Humans.J Cancer. 2020 Jan 17;11(7):1737-1750. doi: 10.7150/jca.37282. eCollection 2020. J Cancer. 2020. PMID: 32194785 Free PMC article. Review.
-
Heat shock protein 90 inhibitors repress latent membrane protein 1 (LMP1) expression and proliferation of Epstein-Barr virus-positive natural killer cell lymphoma.PLoS One. 2013 May 3;8(5):e63566. doi: 10.1371/journal.pone.0063566. Print 2013. PLoS One. 2013. PMID: 23658841 Free PMC article.
References
-
- Ardila-Osorio H., Clausse,B., Mishal,Z., Wiels,J., Tursz,T. and Busson,P. (1999) Evidence of LMP1–TRAF3 interactions in glycosphingolipid-rich complexes of lymphoblastoid and nasopharyngeal carcinoma cells. Int. J. Cancer, 81, 645–649. - PubMed
-
- Barker V., Middleton,G., Davey,F. and Davies,A.M. (2001) TNFα contributes to the death of NGF-dependent neurons during development. Nat. Neurosci., 4, 1194–1198. - PubMed
-
- Berberich I., Shu,G.L. and Clark,E.A. (1994) Cross-linking CD40 on B cells rapidly activates nuclear factor-κB. J. Immunol., 153, 4357–4366. - PubMed
-
- Coffin W.F. III, Erickson,K.D., Hoedt-Miller,M. and Martin,J.M. (2001) The cytoplasmic amino-terminus of the latent membrane protein-1 of Epstein–Barr virus: relationship between transmembrane orientation and effector functions of the carboxy-terminus and transmembrane domain. Oncogene, 20, 5313–5330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
