

Review
doi: 10.1136/heart.89.7.806.
Development of the heart: (1) formation of the cardiac chambers and arterial trunks
Affiliations
- PMID: 12807866
- PMCID: PMC1767747
- DOI: 10.1136/heart.89.7.806
Item in Clipboard
Review
Development of the heart: (1) formation of the cardiac chambers and arterial trunks
Heart.
2003 Jul.
No abstract available
Figures
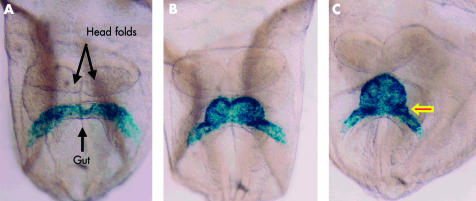
Ventral views of mouse embryos having four, six, and eight somites, respectively (panels A–C), representing the period from 7 ½12 to 8 days after fertilisation, in which the myocardium has been labelled blue using a reporter transgene for myosin light chain. The arrowed bulge in panel C is fated to contribute to the atrioventricular canal.
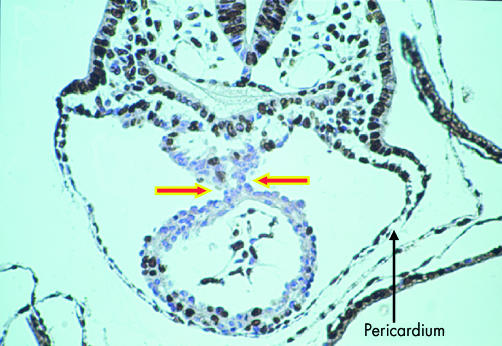
This cross section is through the ventricular part of the heart tube of a mouse embryo with eight somites. At this stage, the tube is supported within the pericardial cavity by a mesocardium (arrows). The tissues have been incorporated with bromodeoxyuridine, which identifies dividing cells, labelled brown. The non-dividing cells are counterstained blue.
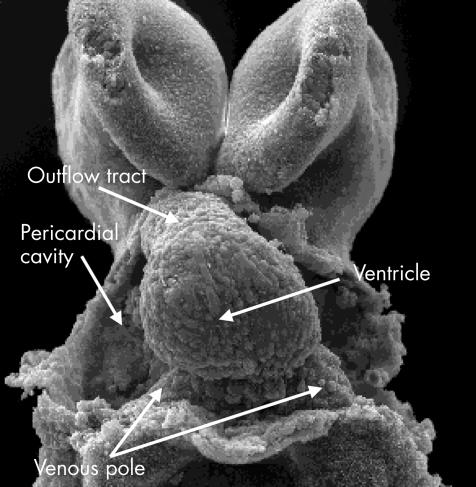
This scanning electron micrograph, viewed from the front in a mouse with 11 somites, shows the stage of the “straight heart tube”. Note that the tube is composed primarily of the ventricle, which will become the definitive left ventricle. As yet, there is no formation of an atrial component, and the outlet is similarly rudimentary.
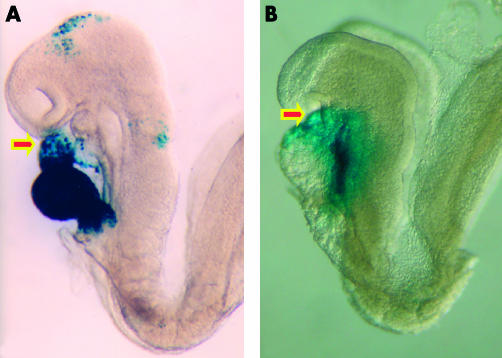
These embryos are photographed in left lateral view. The embryo shown in panel A carries a transgenic marker for myosin light chain, and shows staining throughout the derivatives of the primary heart field. Note that the outflow region (arrowed) is only partly stained. The embryo shown in panel B carries a transgenic marker for fibroblast growth factor 10. This marks the initial site of the contributions made to the developing heart from the secondary heart field, staining the outflow tract (arrowed) and developing right ventricle (see also fig 5).
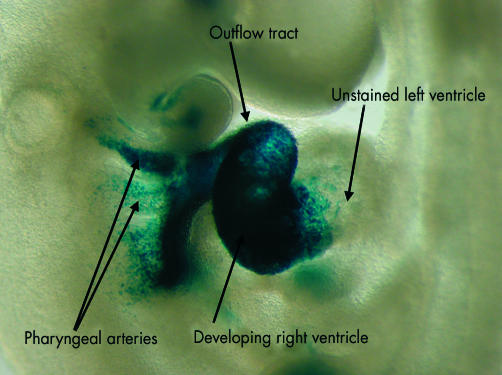
In this embryo, the cells from the secondary heart field, shown in fig 4B, are labelled with a reporter transgene for fibroblast growth factor 10. As can be seen, the cells from the secondary field populate the pharyngeal arches, the outflow tract, and the developing right ventricle. They do not, however, extend beyond the interventricular groove to enter the left ventricle.
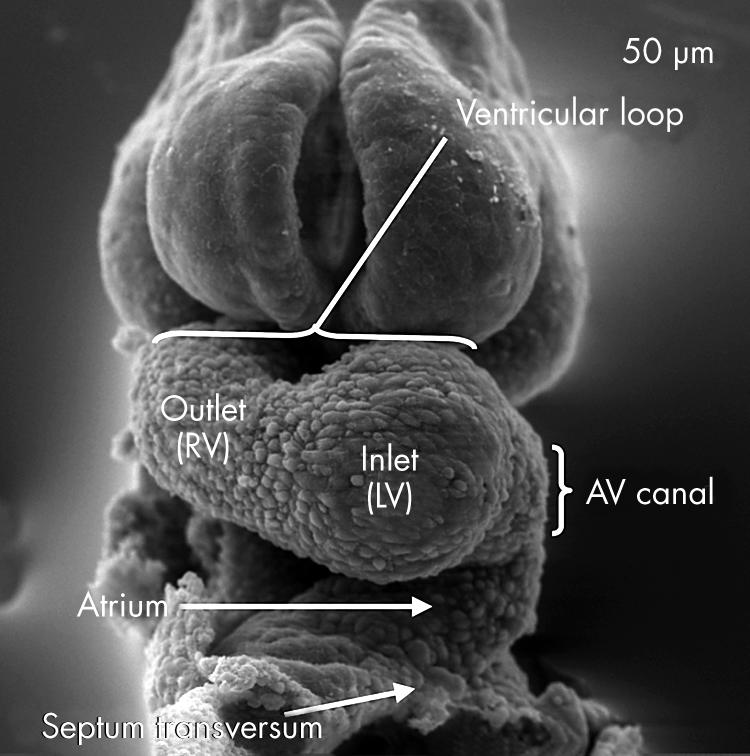
In this scanning electron micrograph, from a mouse embryo with 12 somites, the ventricular component of the heart tube has looped to the right. It now possesses two segments, which develop in sequence. The apical part of the left ventricle (LV) will balloon out from the inlet part of the loop, while the apical part of the right ventricle (RV) will grow from the outlet part. Note also that there is now an atrioventricular canal formed between the ventricular loop and the developing atrial segment.
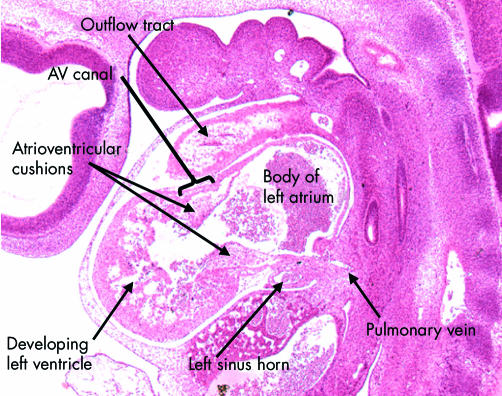
This sagittal section is from a human embryo at Carnegie stage 14. It shows the developing left ventricle in communication with the atrial component of the primary heart tube through the atrioventricular canal, which at this stage has considerable length. Note the presence of the atrioventricular endocardial cushions on the luminal aspect of the canal. Note also the location of the pulmonary vein, which at this stage opens inferiorly to the primary atrial component.
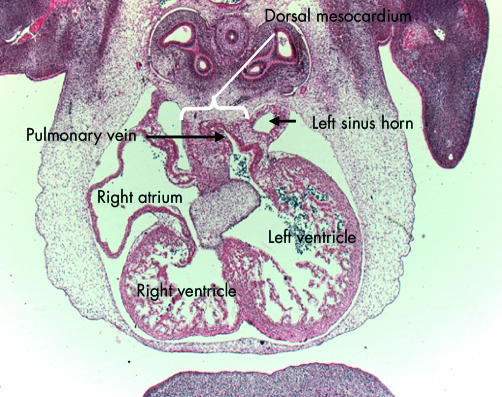
This section, cut in the frontal plane, in a human embryo at Carnegie stage 16, shows the entry of the pulmonary vein to the inferior aspect of the left side of the primary atrial component. Note that the vein has entered through the dorsal mesocardium, and note its relation to the left sinus horn, which will persist as the coronary sinus.

This scanning electron micrograph shows a dissection of the atrial chambers in a mouse embryo with 43 somites, in the second half of the 11th day of development. The systemic venous sinus is now enclosed by the venous valves, and is incorporated within the right side of the primary atrial component. The pulmonary vein opens as a solitary channel to the inferior part of the left side of the primary atrium, entering between the ridges representing the remnants of the dorsal mesocardium. Note the primary atrial septum, which is growing down from the roof of the primary atrium.
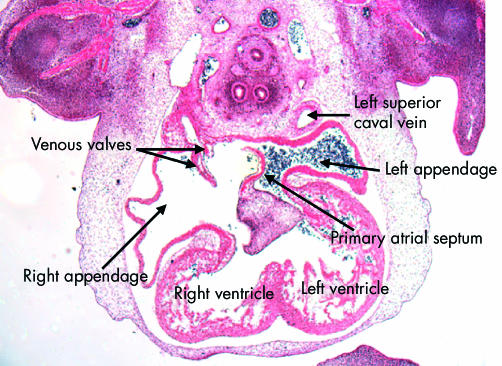
This section, in the frontal plane, is from the human embryo at Carnegie stage 16. It shows the difference, from the outset, in the arrangement of the atrial appendages. The left appendage arises superiorly from a narrow neck in the left side of the primary atrial component. The right appendage, in contrast, is much broader, and is separated by the systemic venous sinus, enclosed within the venous valves, from the atrial septum. Note that the primary septum has now grown down and fused with the superior atrioventricular endocardial cushion. Note also that the left superior caval vein possesses its own walls outside the developing left atrium.
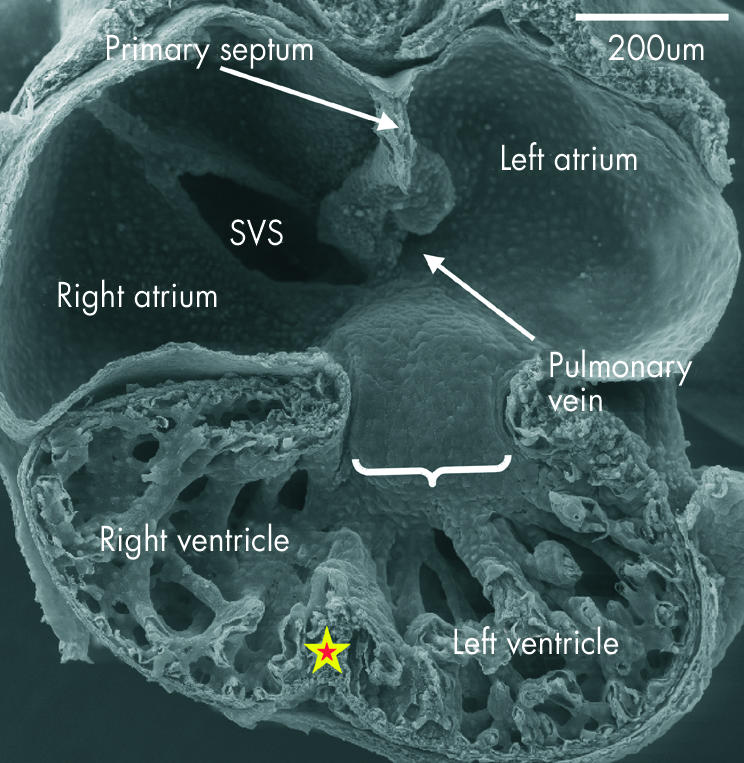
This scanning electron micrograph shows the posterior half of a mouse embryo with 40 somites transacted in the frontal plane. The systemic venous sinus (SVS) is incorporated within the right side of the primary atrium, and the ridges marking the site of the dorsal mesocardium are seen inferiorly in the left side. The pulmonary vein opens between these ridges (see fig 9). The atrioventricular canal at this stage, shown by the bracket, with the inferior cushion occupying most of its wall, is connected through the larger part of its circumference with the inlet part of the ventricular component of the primary heart tube, from which is ballooning the apical part of the developing left ventricle. Note the small size, at this stage, of the developing right and left atrioventricular orifices. The apical part of the developing right ventricle is ballooning from the outlet part of the primary tube, and the primordium of the muscular ventricular septum, shown by the star, is seen between the two pouches.
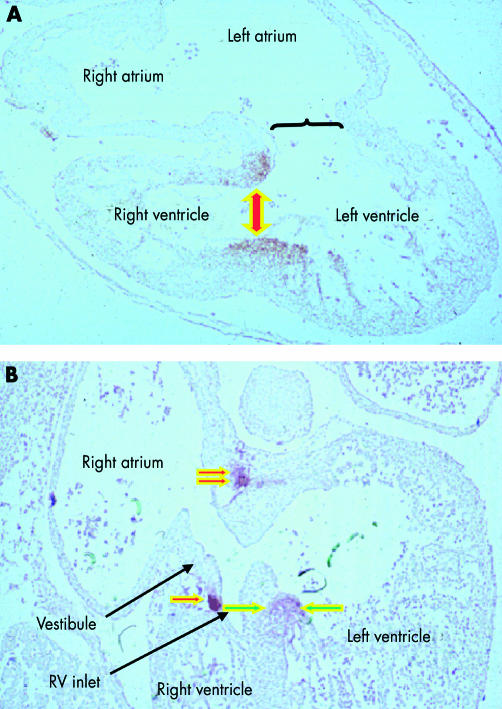
These sections, from human embryos, show the fate of the primary interventricular foramen. This is shown in the upper panel, from an embryo at Carnegie stage 15, in which the larger circumference of the atrioventricular canal is supported by the inlet component of the ventricular loop, from which has ballooned the apical part of the left ventricle. The muscle surrounding the interventricular foramen (shown by the double headed arrow) is demarcated by an antibody to the nodose ganglion of the chick. The lower panel shows the stage subsequent to expansion of the atrioventricular canal, representing Carnegie stage 16 or 17. The right side of the atrioventricular canal itself has now been incorporated into the right atrium as the vestibule of the forming tricuspid valve. The location of the tissue staining positive for the antibody (single red arrow) shows that the inlet of the right ventricle has been derived from the outlet component of the ventricular part of the primary heart tube. Note that the tissue demarcating the location of the initial interventricular foramen retains its location on the crest of the muscular septum (between green arrows), but is also extending behind the outflow tract (double red arrow) concomitant with the transfer of the subaortic outlet to the developing left ventricle.
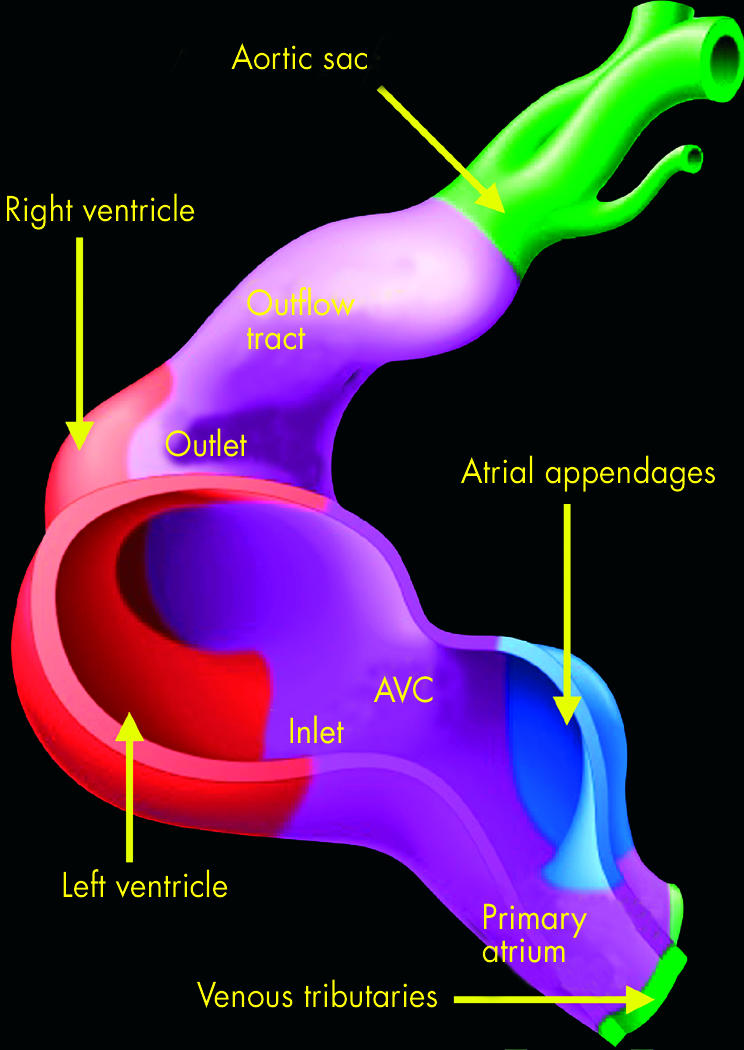
This illustration shows the origin of the components of the developing atriums and ventricles. The myocardium of the primary heart tube is shown in purple, and makes up the primary atrium, the atrioventricular canal (AVC), the inlet and outlet components of the ventricular loop, and the outflow tract. Shown in green are the systemic venous tributaries, which are eventually incorporated within the right atrium, and the aortic sac with its arterial branches. The pulmonary vein is not shown, this being a new development appearing concomitant with the formation of the lungs. The atrial appendages, shown in blue, balloon in parallel from the primary atrial component of the heart tube. The apical parts of the ventricles, in contrast, balloon in series from the primary tube, with the apical part of the left ventricle growing from the inlet component, and the apical part of the right ventricle from the outlet component.
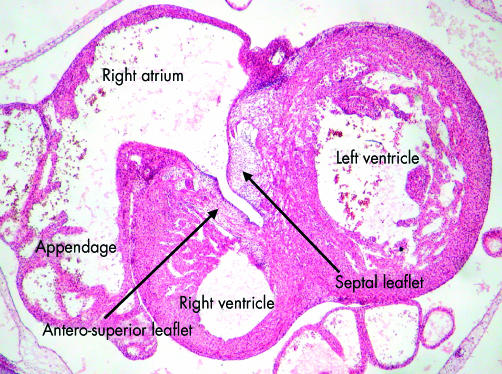
This section, from a human embryo at Carnegie stage 20, just subsequent to the completion of ventricular septation, shows how the leaflets of the tricuspid valve are delaminating within the part of the primary heart tube from which ballooned the apical component of the right ventricle. As the leaflets delaminate from the superficial layers of ventricular myocardium, they incorporate the endocardial cushions within their substance, subsequently being transformed into fibrous tissue.
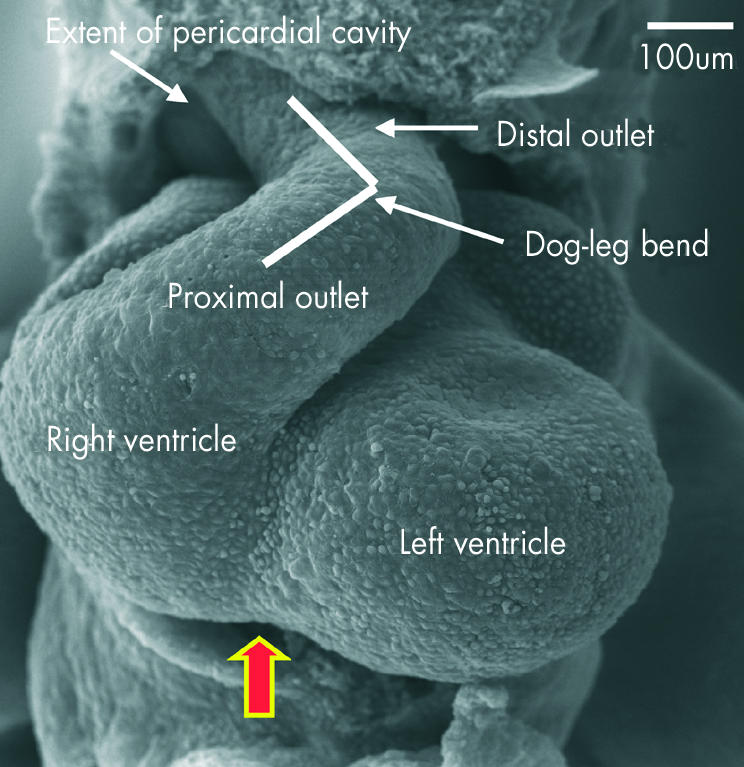
This scanning electron micrograph, from a mouse with 40 somites, shows how the outflow tract, now supported by the developing right ventricle, has bent into two discrete components separated by a dog-leg bend. The distal part extends to the border of the pericardial cavity, where it becomes continuous with the aortic sac. The arrow shows the interventricular groove.
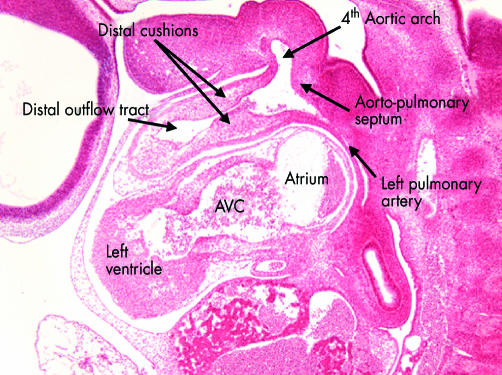
This sagittal section is from the human embryo at Carnegie stage 14 also illustrated in fig 7. This section is taken through the distal part of the outflow tract, and shows the distal cushions that will divide this part into the intrapericardial portions of the aorta and pulmonary trunk (see figs 17 and 18). Note the location of the aortopulmonary septum within the aortic sac, separating the origins of the fourth and sixth arches. In this section the left pulmonary artery is seen arising from the artery of the sixth arch. AVC, atrioventricular canal.
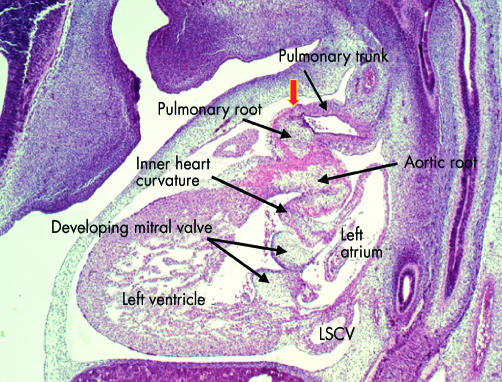
This sagittal section is from a human embryo at Carnegie stage 18, just before the completion of cardiac septation. Note that the distal outflow tract has now separated into the intrapericardial components of the pulmonary trunk, which is sectioned, and the aorta, which cannot be seen in this section. The arterial roots at this stage, however, are still encased within the muscular wall of the proximal outflow tract. The sinutubular junction, shown by the arrow, is formed at the site of the initial dog-leg bend (see fig 15). Note the cushions fusing to form the aortic leaflet of the mitral valve, which at this stage is still separated by the musculature of the inner heart curvature from the developing leaflets of the aortic valve. Note also the left superior caval vein (LSCV) now incorporated within its own wall within the left atrioventricular groove.
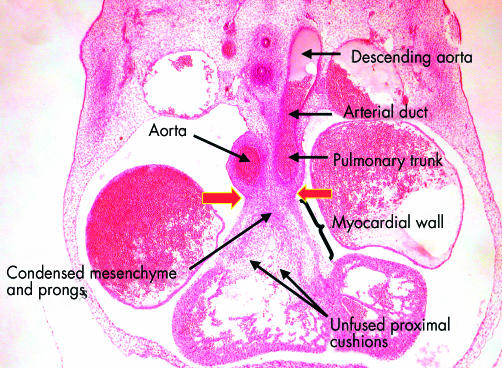
This frontal section, from a human embryo at Carnegie stage 16, shows the distinction between the proximal and distal parts of the outflow tract. The distal part has been separated by the distal cushions into the intrapericardial portions of the aorta and the pulmonary trunk, the walls of which now possess an arterial phenotype. Note the continuation of the pulmonary trunk through the arterial duct, derived from the sixth arch, to the descending aorta. The proximal outflow tract, in contrast, is still enclosed within muscular walls. The arrows show the site of the developing sinutubular junction, formed at the initial dog-leg bend. Note that the proximal cushions are fused distally, with the site of fusion marked by condensed mesenchyme. The more proximal parts of the cushions, however, penetrated by prongs from the condensed mesenchymal mass, remain unfused. Completion of fusion, followed by muscularisation, will convert this area into the supraventricular crest of the right ventricle.
References
-
- Lints TJ, Parsons LM, Hartley L, et al. Nkx-2.5: a novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993;119:419–31. - PubMed
-
- de la Cruz MV, Sanchez-Gómez C, Cayre R. The developmental components of ventricles: their significance in congenital malformations. Cardiol Young 1991;1:123–128. ▸ Maria Victoria de la Cruz was one of the first embryologists to recognise the fact that all parts of the developing heart were not already present at the “straight tube” stage. This paper is a review of her earlier works, and puts the embryological studies into the context of the morphology of congenital cardiac malformations.
-
- Black BL, Olson EN. Control of cardiac development by the MEF2 family of transcription factors. In: Harvey RP, Rosenthal N, eds. Heart development. London: Academic Press, 1999:131–42. ▸ A good chapter in an excellent textbook. As with many of the other chapters, however, the value of the observations concerning advances in molecular biology are devalued to some extent by slavish adherence to concepts of “armchair embryology”.
-
- Knauth A, McCarthy KP, Webb S, et al. Interatrial communication through the mouth of the coronary sinus defect. Cardiol Young 2002;12:364–72. ▸ An important study showing that, from the start of its development, the coronary sinus, derived from the left sinus horn, possessed its own walls discrete from those of the left atrium. - PubMed
-
- Webb S, Kanani M, Anderson RH, et al. Development of the human pulmonary vein and its incorporation in the morphologically left atrium. Cardiol Young 2001;11:632–42. ▸ Conventional wisdom has suggested that the pulmonary vein is derived from the embryonic systemic venous sinus, or “sinus venosus”. This study showed that there is no evidence to support this notion, since the incorporation of the pulmonary vein to the left atrium is a late event in cardiac development. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources