

The murine Nck SH2/SH3 adaptors are important for the development of mesoderm-derived embryonic structures and for regulating the cellular actin network
- PMID: 12808099
- PMCID: PMC164855
- DOI: 10.1128/MCB.23.13.4586-4597.2003
The murine Nck SH2/SH3 adaptors are important for the development of mesoderm-derived embryonic structures and for regulating the cellular actin network
Abstract
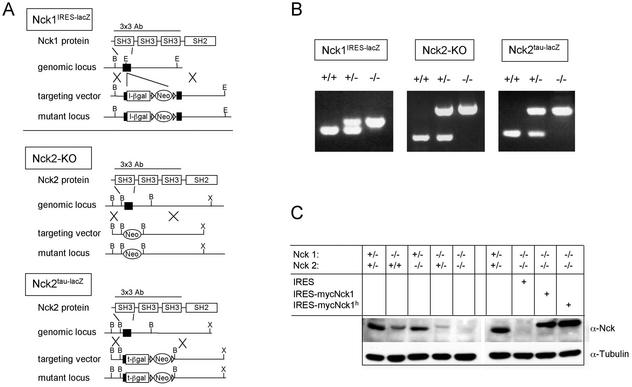
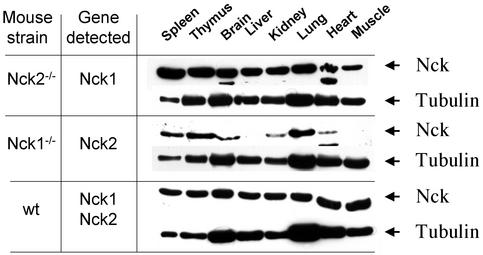
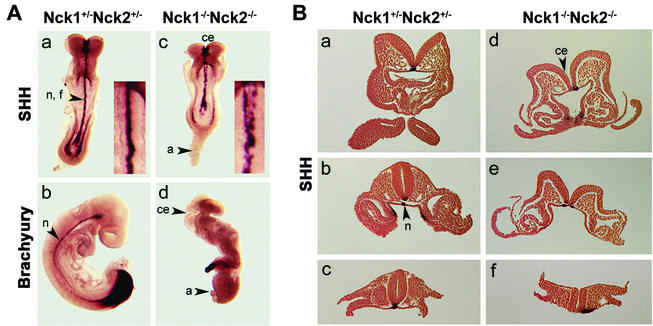
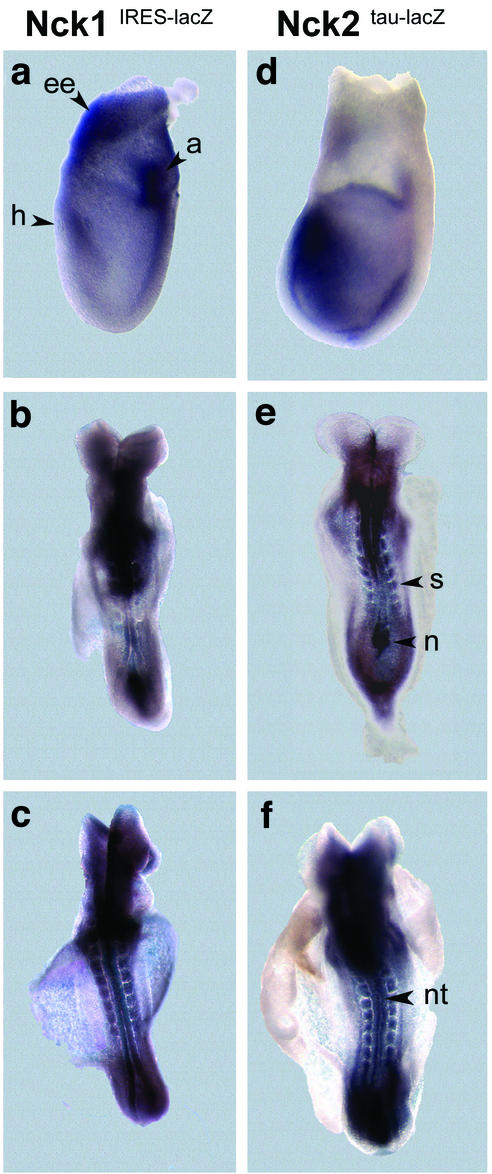
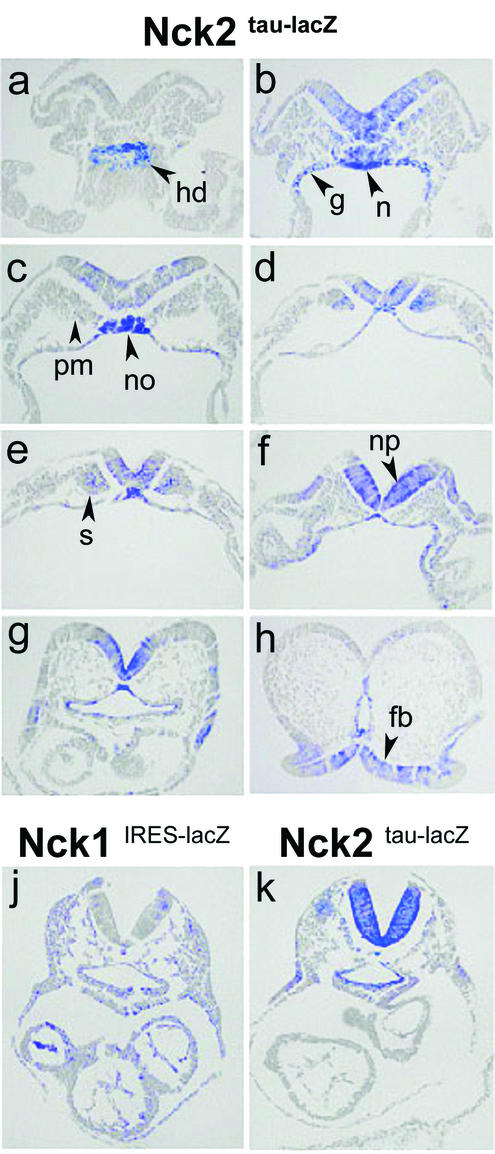
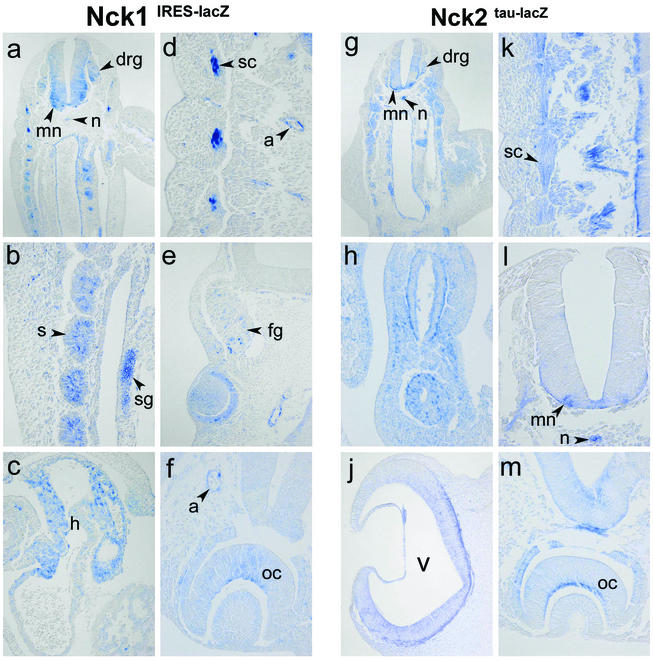
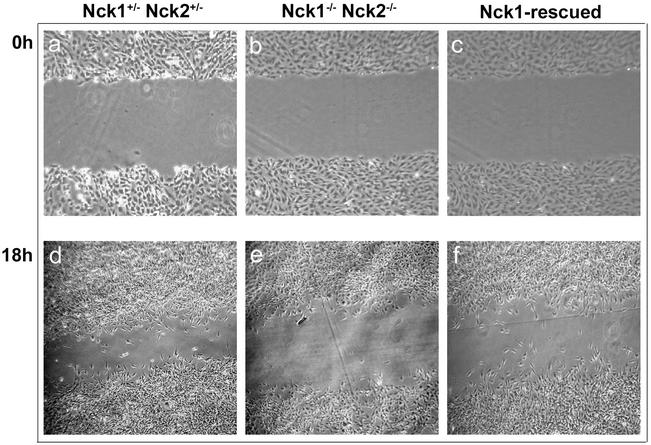
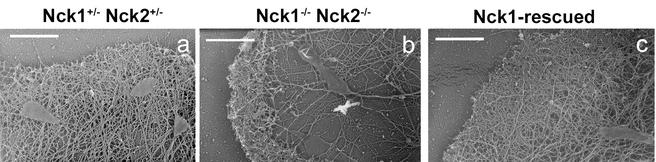
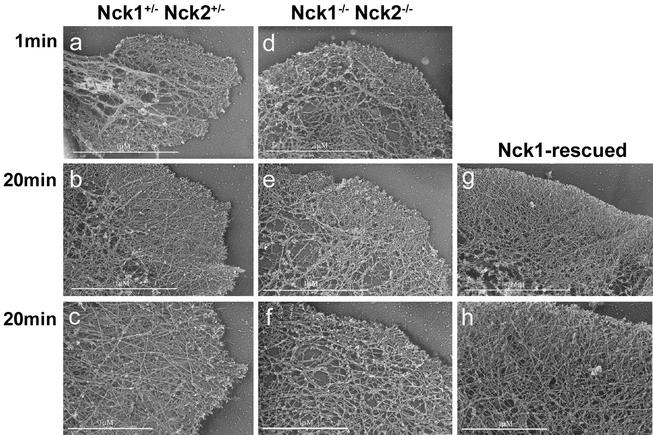
Mammalian Nck1 and Nck2 are closely related adaptor proteins that possess three SH3 domains, followed by an SH2 domain, and are implicated in coupling phosphotyrosine signals to polypeptides that regulate the actin cytoskeleton. However, the in vivo functions of Nck1 and Nck2 have not been defined. We have mutated the murine Nck1 and Nck2 genes and incorporated beta-galactosidase reporters into the mutant loci. In mouse embryos, the two Nck genes have broad and overlapping expression patterns. They are functionally redundant in the sense that mice deficient for either Nck1 or Nck2 are viable, whereas inactivation of both Nck1 and Nck2 results in profound defects in mesoderm-derived notochord and embryonic lethality at embryonic day 9.5. Fibroblast cell lines derived from Nck1(-/-) Nck2(-/-) embryos have defects in cell motility and in the organization of the lamellipodial actin network. These data suggest that the Nck SH2/SH3 adaptors have important functions in the development of mesodermal structures during embryogenesis, potentially linked to a role in cell movement and cytoskeletal organization.
Figures
References
-
- Anton, I. M., W. Lu, B. J. Mayer, N. Ramesh, and R. S. Geha. 1998. The Wiskott-Aldrich syndrome protein-interacting protein (WIP) binds to the adaptor protein Nck. J. Biol. Chem. 273:20992-20995. - PubMed
-
- Beddington, R. S. 1994. Induction of a second neural axis by the mouse node. Development 120:613-620. - PubMed
-
- Bokoch, G. M., Y. Wang, B. P. Bohl, M. A. Sells, L. A. Quilliam, and U. G. Knaus. 1996. Interaction of the Nck adapter protein with p21-activated kinase (PAK1). J. Biol. Chem. 271:25746-25749. - PubMed
-
- Brand-Saberi, B., and B. Christ. 2000. Evolution and development of distinct cell lineages derived from somites. Curr. Top. Dev. Biol. 48:1-42. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases