

The Bcr kinase downregulates Ras signaling by phosphorylating AF-6 and binding to its PDZ domain
- PMID: 12808105
- PMCID: PMC164848
- DOI: 10.1128/MCB.23.13.4663-4672.2003
The Bcr kinase downregulates Ras signaling by phosphorylating AF-6 and binding to its PDZ domain
Abstract
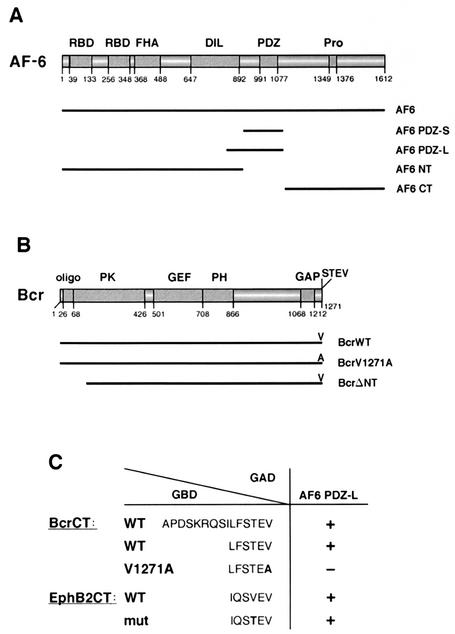
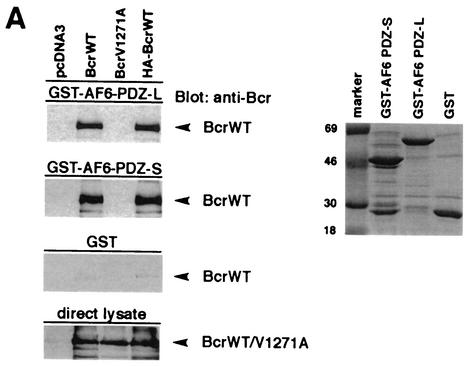
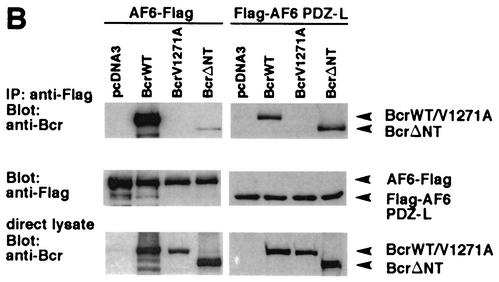
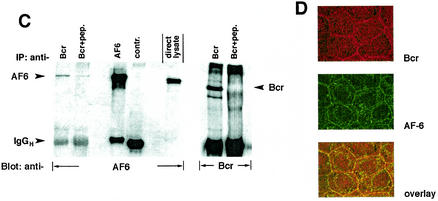
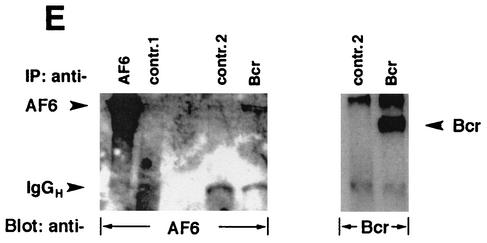
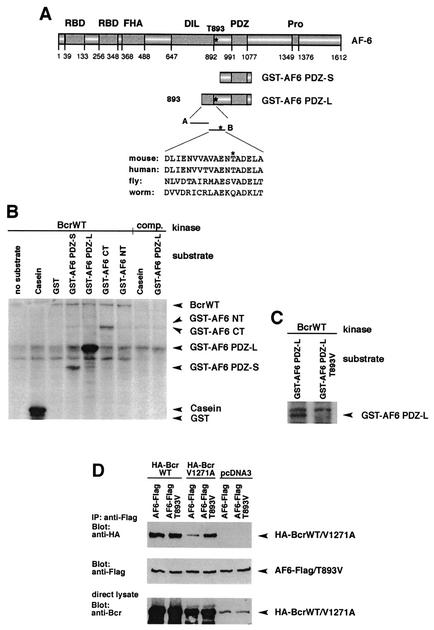
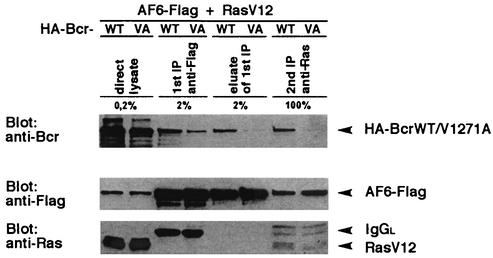
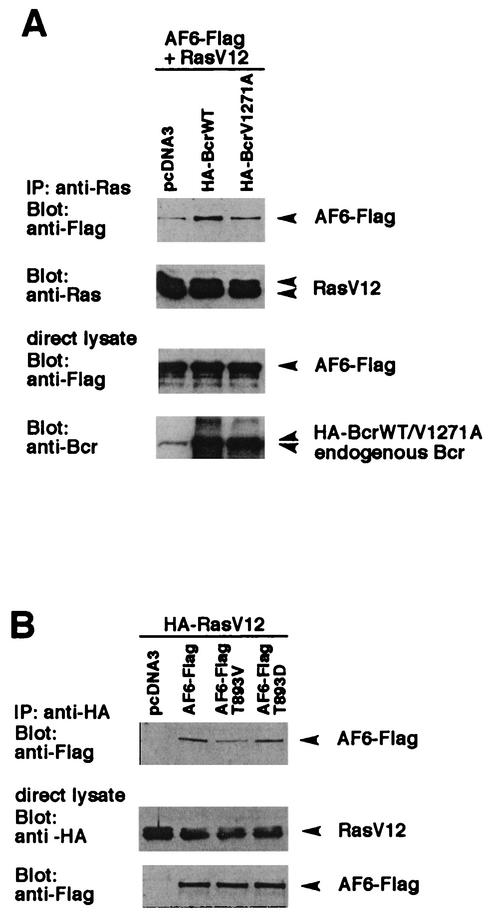
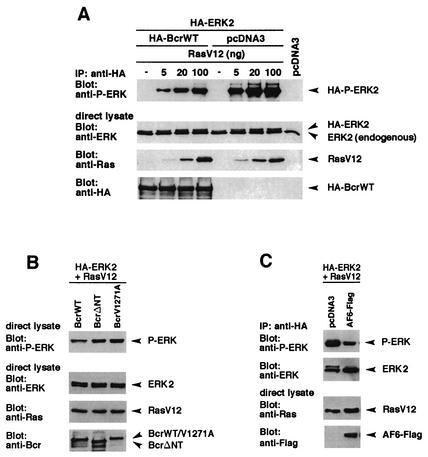
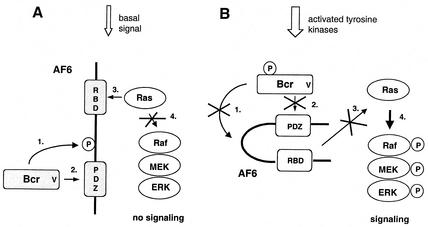
The protein kinase Bcr is a negative regulator of cell proliferation and oncogenic transformation. We identified Bcr as a ligand for the PDZ domain of the cell junction and Ras-interacting protein AF-6. The Bcr kinase phosphorylates AF-6, which subsequently allows efficient binding of Bcr to AF-6, showing that the Bcr kinase is a regulator of the PDZ domain-ligand interaction. Bcr and AF-6 colocalize in epithelial cells at the plasma membrane. In addition, Bcr, AF-6, and Ras form a trimeric complex. Bcr increases the affinity of AF-6 to Ras, and a mutant of AF-6 that lacks a specific phosphorylation site for Bcr shows a reduced binding to Ras. Wild-type Bcr, but not Bcr mutants defective in binding to AF-6, interferes with the Ras-dependent stimulation of the Raf/MEK/ERK pathway. Since AF-6 binds to Bcr via its PDZ domain and to Ras via its Ras-binding domain, we propose that AF-6 functions as a scaffold-like protein that links Bcr and Ras to cellular junctions. We suggest that this trimeric complex is involved in downregulation of Ras-mediated signaling at sites of cell-cell contact to maintain cells in a nonproliferating state.
Figures
Similar articles
-
Solution structure and backbone dynamics of the AF-6 PDZ domain/Bcr peptide complex.Protein Sci. 2007 Jun;16(6):1053-62. doi: 10.1110/ps.062440607. Epub 2007 May 1. Protein Sci. 2007. PMID: 17473018 Free PMC article.
-
Solution structure of AF-6 PDZ domain and its interaction with the C-terminal peptides from Neurexin and Bcr.J Biol Chem. 2005 Apr 8;280(14):13841-7. doi: 10.1074/jbc.M411065200. Epub 2005 Jan 31. J Biol Chem. 2005. PMID: 15684424
-
Signaling complexes and protein-protein interactions involved in the activation of the Ras and phosphatidylinositol 3-kinase pathways by the c-Ret receptor tyrosine kinase.J Biol Chem. 2000 Dec 15;275(50):39159-66. doi: 10.1074/jbc.M006908200. J Biol Chem. 2000. PMID: 10995764
-
Meaningful relationships: the regulation of the Ras/Raf/MEK/ERK pathway by protein interactions.Biochem J. 2000 Oct 15;351 Pt 2(Pt 2):289-305. Biochem J. 2000. PMID: 11023813 Free PMC article. Review.
-
Interactions between Ras and Raf: key regulatory proteins in cellular transformation.Mol Reprod Dev. 1995 Dec;42(4):493-9. doi: 10.1002/mrd.1080420418. Mol Reprod Dev. 1995. PMID: 8607981 Review.
Cited by
-
The PDZ protein Canoe/AF-6 links Ras-MAPK, Notch and Wingless/Wnt signaling pathways by directly interacting with Ras, Notch and Dishevelled.PLoS One. 2006 Dec 20;1(1):e66. doi: 10.1371/journal.pone.0000066. PLoS One. 2006. PMID: 17183697 Free PMC article.
-
Regulation of c-Src by binding to the PDZ domain of AF-6.EMBO J. 2007 Jun 6;26(11):2633-44. doi: 10.1038/sj.emboj.7601706. Epub 2007 May 10. EMBO J. 2007. PMID: 17491594 Free PMC article.
-
Loss of polarity protein AF6 promotes pancreatic cancer metastasis by inducing Snail expression.Nat Commun. 2015 May 26;6:7184. doi: 10.1038/ncomms8184. Nat Commun. 2015. PMID: 26013125
-
Modelling Cooperative Tumorigenesis in Drosophila.Biomed Res Int. 2018 Mar 6;2018:4258387. doi: 10.1155/2018/4258387. eCollection 2018. Biomed Res Int. 2018. PMID: 29693007 Free PMC article. Review.
-
The nuclear RhoA exchange factor Net1 interacts with proteins of the Dlg family, affects their localization, and influences their tumor suppressor activity.Mol Cell Biol. 2007 Dec;27(24):8683-97. doi: 10.1128/MCB.00157-07. Epub 2007 Oct 15. Mol Cell Biol. 2007. PMID: 17938206 Free PMC article.
References
-
- Boguski, M. S., and F. McCormick. 1993. Proteins regulating Ras and its relatives. Nature 366:643-654. - PubMed
-
- Buchert, M., S. Schneider, V. Meskenaite, M. T. Adams, E. Canaani, T. Baechi, K. Moelling, and C. M. Hovens. 1999. The junction-associated protein AF-6 interacts and clusters with specific Eph receptor tyrosine kinases at specialized sites of cell-cell contact in the brain. J. Cell Biol. 144:361-371. - PMC - PubMed
-
- Durfee, T., K. Becherer, P. L. Chen, S. H. Yeh, Y. Yang, A. E. Kilburn, W. H. Lee, and S. J. Elledge. 1993. The retinoblastoma protein associates with the protein phosphatase type 1 catalytic subunit. Genes Dev. 7:555-569. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous