

Multistep nature of microvascular recruitment of ex vivo-expanded embryonic endothelial progenitor cells during tumor angiogenesis
- PMID: 12810693
- PMCID: PMC2193947
- DOI: 10.1084/jem.20021659
Multistep nature of microvascular recruitment of ex vivo-expanded embryonic endothelial progenitor cells during tumor angiogenesis
Abstract
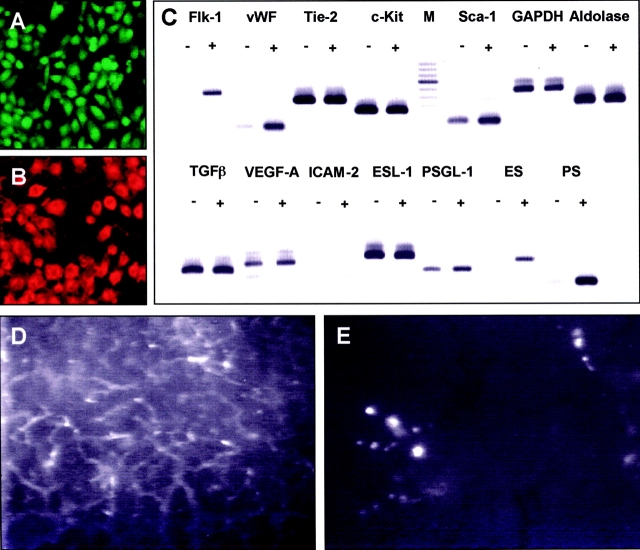
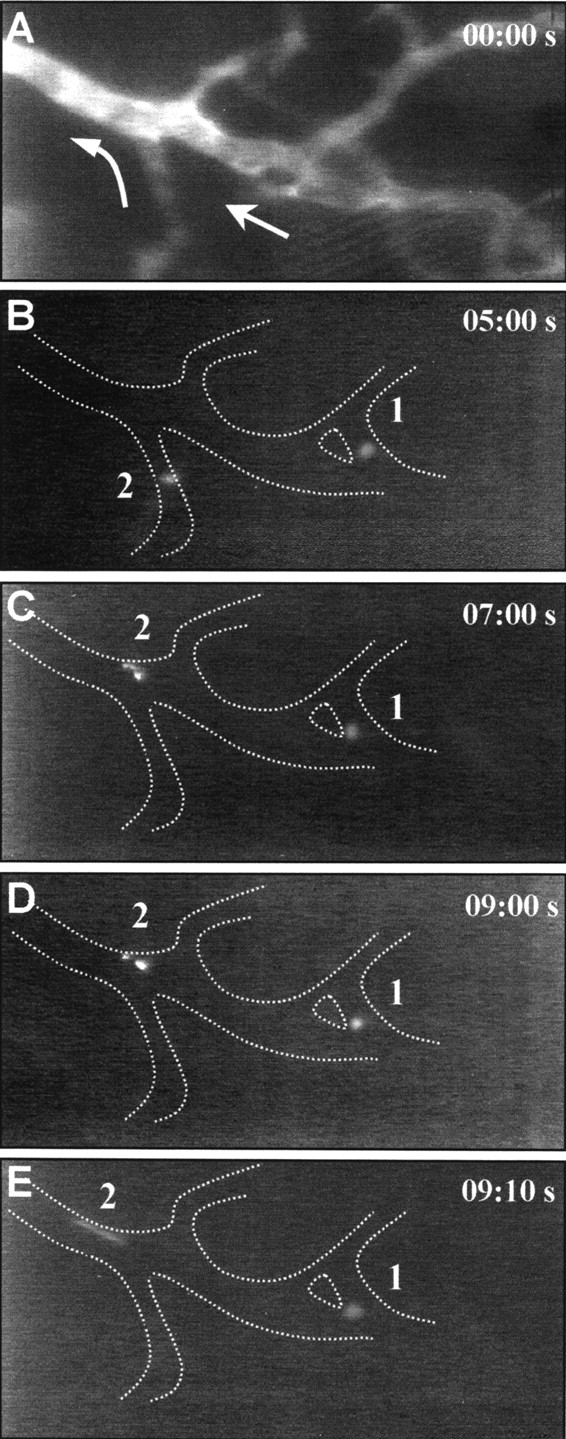
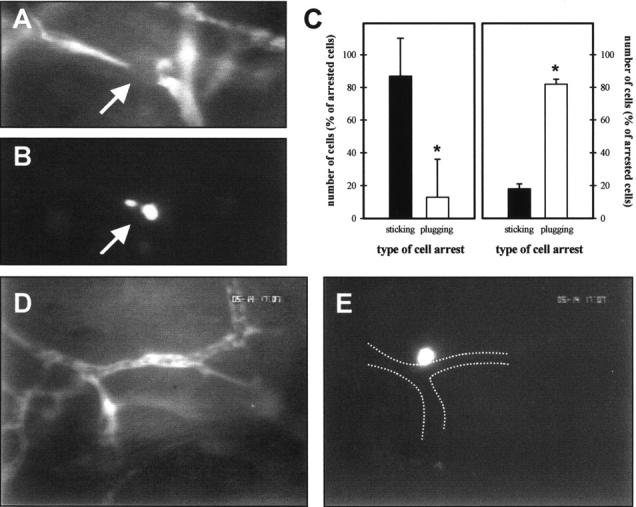
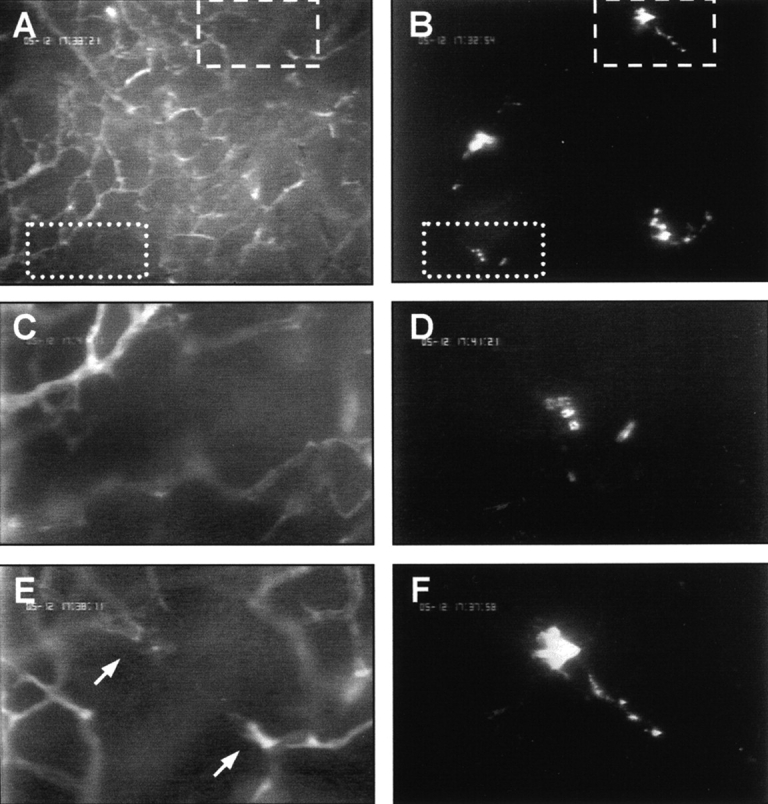
Tissue neovascularization involves recruitment of circulating endothelial progenitor cells that originate in the bone marrow. Here, we show that a class of embryonic endothelial progenitor cells (Tie-2+, c-Kit+, Sca-1+, and Flk-1-/low), which were isolated at E7.5 of mouse development at the onset of vasculogenesis, retain their ability to contribute to tumor angiogenesis in the adult. Using intravital fluorescence videomicroscopy, we further defined the multistep process of embryonic endothelial progenitor cell (eEPC) homing and incorporation. Circulating eEPCs are specifically arrested in "hot spots" within the tumor microvasculature, extravasate into the interstitium, form multicellular clusters, and incorporate into functional vascular networks. Expression analysis and in vivo blocking experiments provide evidence that the initial cell arrest of eEPC homing is mediated by E- and P-selectin and P-selectin glycoprotein ligand 1. This paper provides the first in vivo insights into the mechanisms of endothelial progenitor cell recruitment and, thus, indicates novel ways to interfere with pathological neovascularization.
Figures
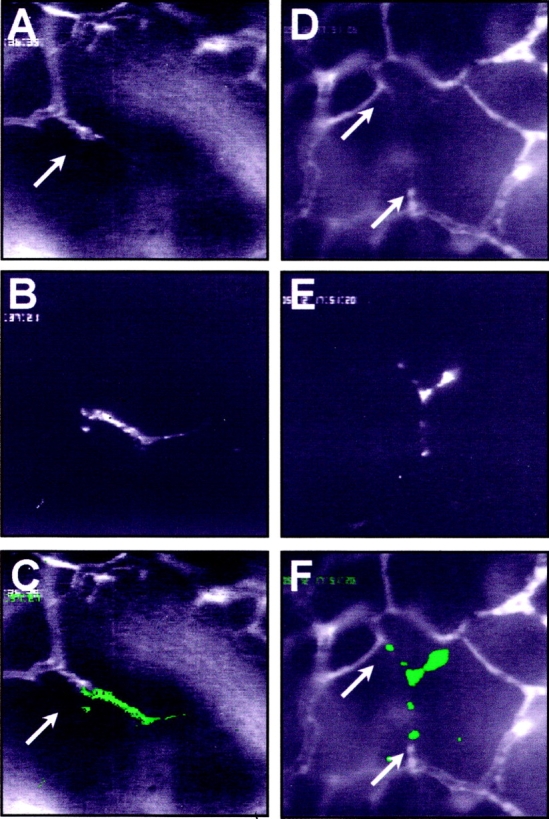
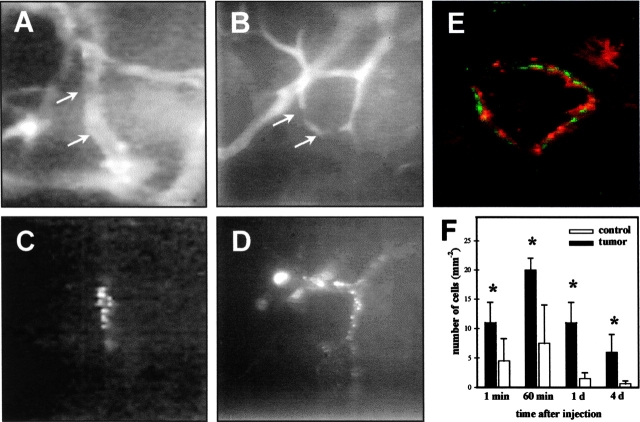
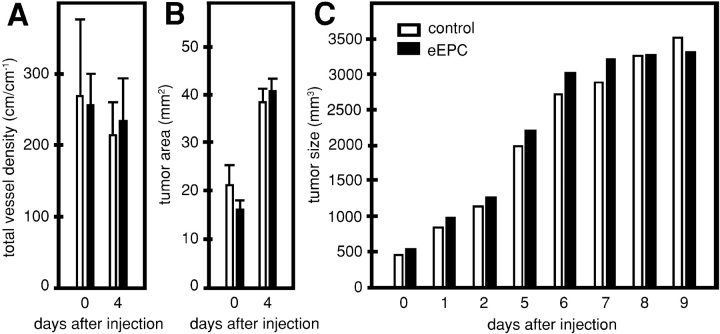
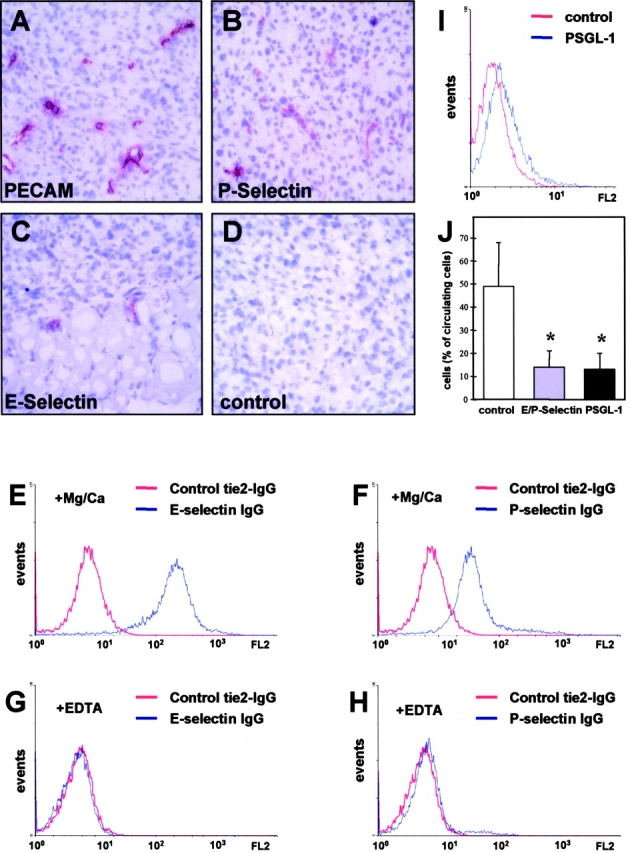
References
-
- Asahara, T., H. Masuda, T. Takahashi, C. Kalka, C. Pastore, M. Silver, M. Kearne, M. Magner, and J.M. Isner. 1999. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ. Res. 85:221–228. - PubMed
-
- Takahashi, T., C. Kalka, H. Masuda, D. Chen, M. Silver, M. Kearney, M. Magner, J.M. Isner, and T. Asahara. 1999. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat. Med. 5:434–438. - PubMed
-
- Kalka, C., H. Masuda, T. Takahashi, R. Gordon, O. Tepper, E. Gravereaux, A. Pieczek, H. Iwaguro, S.I. Hayashi, J.M. Isner, and T. Asahara. 2000. Vascular endothelial growth factor(165) gene transfer augments circulating endothelial progenitor cells in human subjects. Circ. Res. 86:1198–1202. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
