

ADP ribosylation factor 6 is activated and controls membrane delivery during phagocytosis in macrophages
- PMID: 12810696
- PMCID: PMC2172982
- DOI: 10.1083/jcb.200210069
ADP ribosylation factor 6 is activated and controls membrane delivery during phagocytosis in macrophages
Abstract
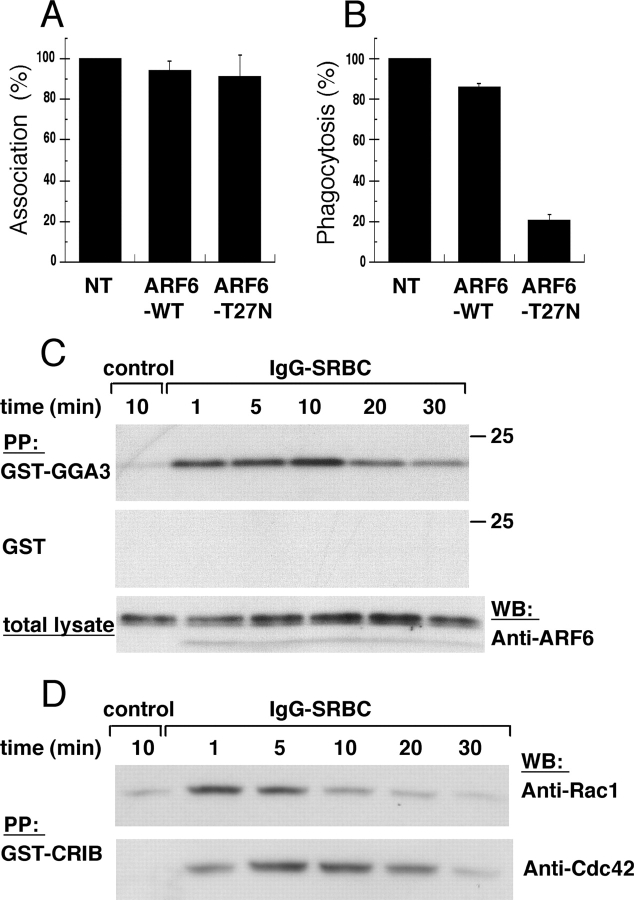
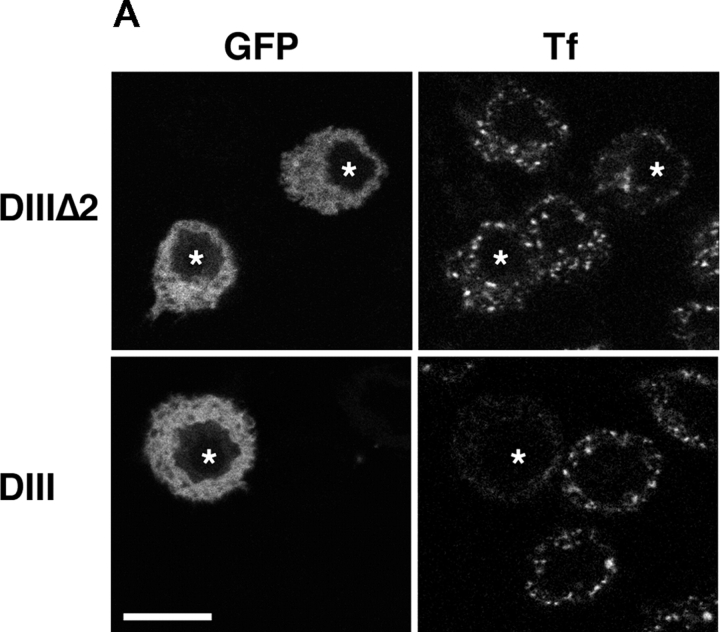
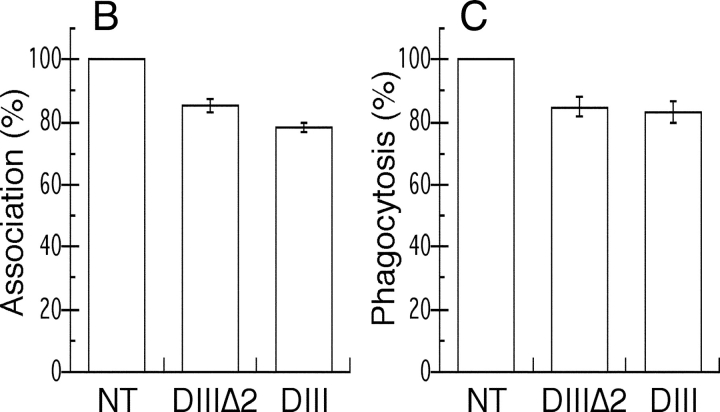
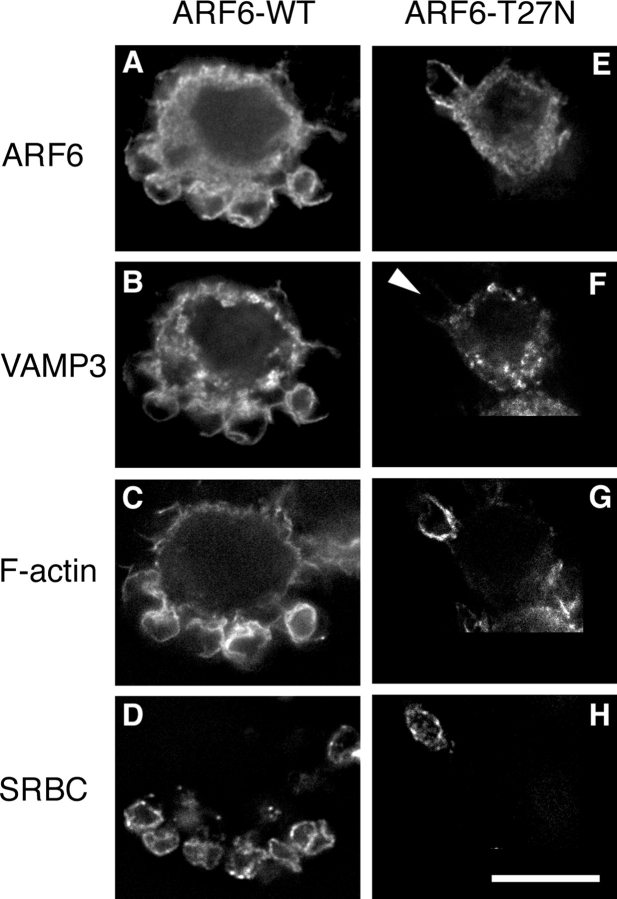
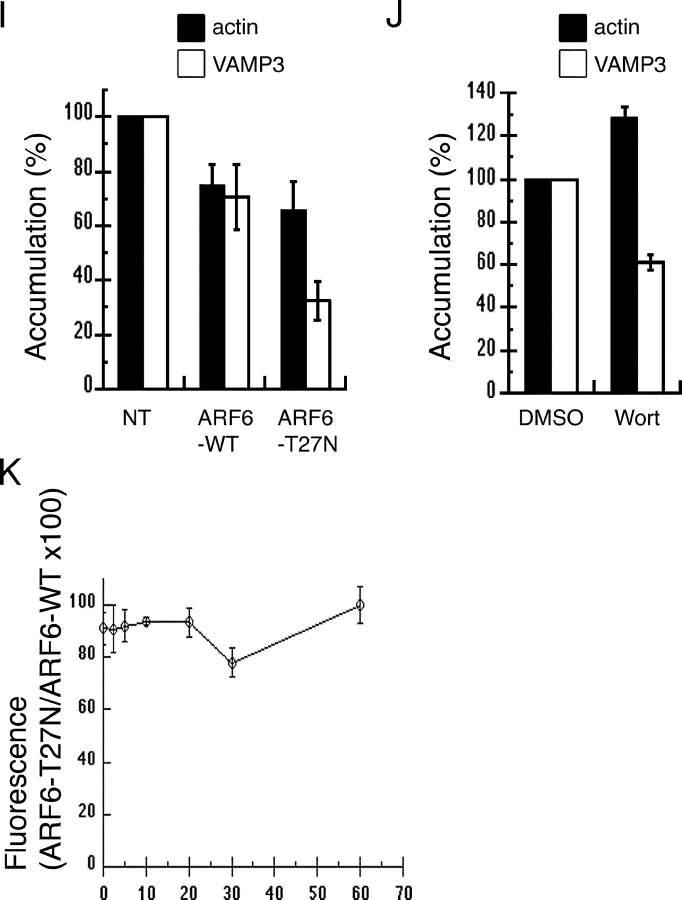
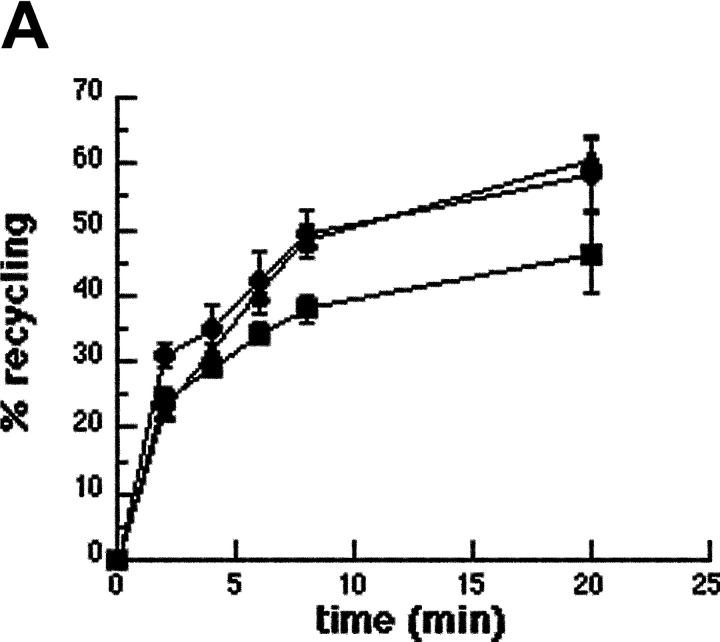
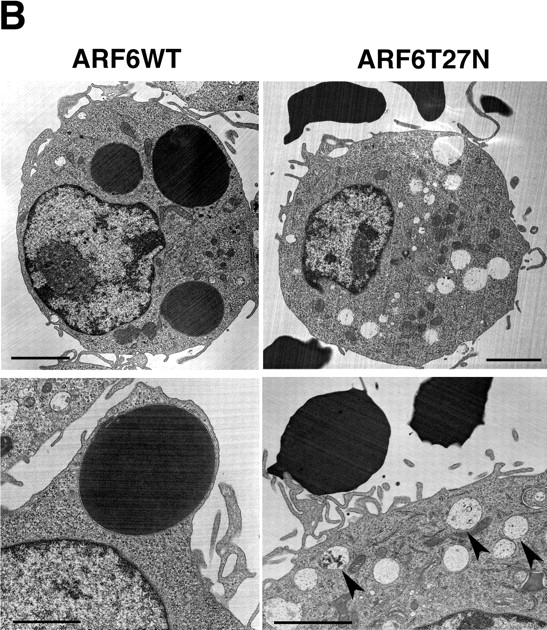
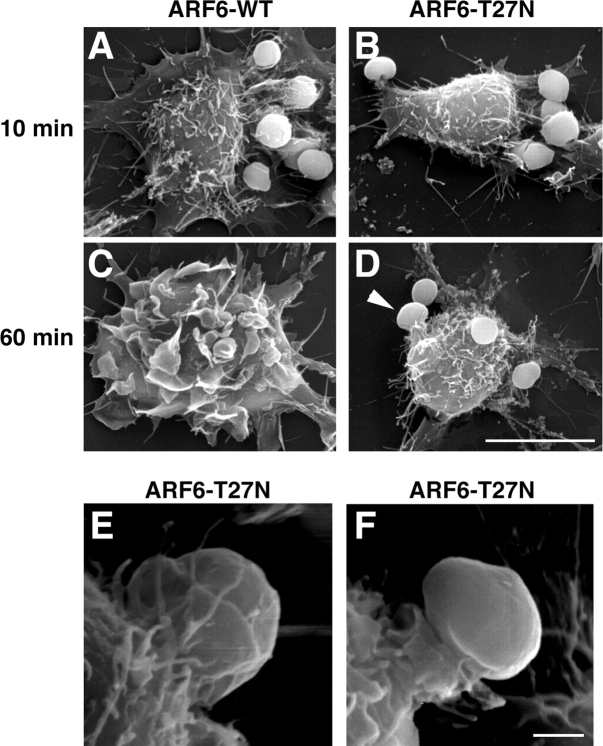
Engulfment of particles by phagocytes is induced by their interaction with specific receptors on the cell surface, which leads to actin polymerization and the extension of membrane protrusions to form a closed phagosome. Membrane delivery from internal pools is considered to play an important role in pseudopod extension during phagocytosis. Here, we report that endogenous ADP ribosylation factor 6 (ARF6), a small GTP-binding protein, undergoes a sharp and transient activation in macrophages when phagocytosis was initiated via receptors for the Fc portion of immunoglobulins (FcRs). A dominant-negative mutant of ARF6 (T27N mutation) dramatically affected FcR-mediated phagocytosis. Expression of ARF6-T27N lead to a reduction in the focal delivery of vesicle-associated membrane protein 3+ endosomal recycling membranes at phagocytosis sites, whereas actin polymerization was unimpaired. This resulted in an early blockade in pseudopod extension and accumulation of intracellular vesicles, as observed by electron microscopy. We conclude that ARF6 is a major regulator of membrane recycling during phagocytosis.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
