

Heterologous immunity provides a potent barrier to transplantation tolerance
- PMID: 12813024
- PMCID: PMC161424
- DOI: 10.1172/JCI17477
Heterologous immunity provides a potent barrier to transplantation tolerance
Abstract
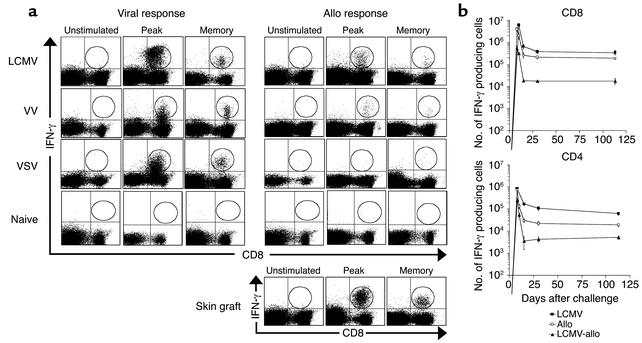
Many strategies have been proposed to induce tolerance to transplanted tissue in rodents; however, few if any have shown equal efficacy when tested in nonhuman primate transplant models. We hypothesized that a critical distinction between specific pathogen-free mice and nonhuman primates or human patients is their acquired immune history. Here, we show that a heterologous immune response--specifically, virally induced alloreactive memory--is a potent barrier to tolerance induction. A critical threshold of memory T cells is needed to promote rejection, and CD8(+) "central" memory T cells are primarily responsible. Finally, treatment with deoxyspergualin, an inhibitor of NF-kappa B translocation, together with costimulation blockade, synergistically impairs memory T cell activation and promotes antigen-specific tolerance of memory. These data offer a potential explanation for the difficulty encountered when inducing tolerance in nonhuman primates and human patients and provide insight into the signaling pathways essential for memory T cell activation and function.
Figures




Comment in
-
Tolerance: of mice and men.J Clin Invest. 2003 Jun;111(12):1819-21. doi: 10.1172/JCI18926. J Clin Invest. 2003. PMID: 12813017 Free PMC article.
References
-
- Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: implications for vaccine development. Nat. Rev. Immunol. 2002;2:251–262. - PubMed
-
- Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. - PubMed
-
- Masopust D, Vezys V, Marzo AL, Lefrancois L. Preferential localization of effector memory cells in nonlymphoid tissue. Science. 2001;291:2413–2417. - PubMed
-
- Reinhardt RL, Khoruts A, Merica R, Zell T, Jenkins MK. Visualizing the generation of memory CD4 T cells in the whole body. Nature. 2001;410:101–105. - PubMed
-
- Welsh RM, Selin LK. No one is naive: the significance of heterologous T-cell immunity. Nat. Rev. Immunol. 2002;2:417–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
