

Host microenvironment in breast cancer development: epithelial-cell-stromal-cell interactions and steroid hormone action in normal and cancerous mammary gland
- PMID: 12817994
- PMCID: PMC165024
- DOI: 10.1186/bcr615
Host microenvironment in breast cancer development: epithelial-cell-stromal-cell interactions and steroid hormone action in normal and cancerous mammary gland
Abstract
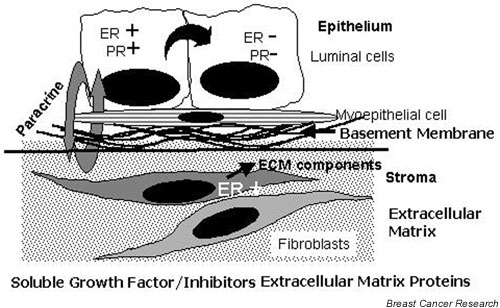
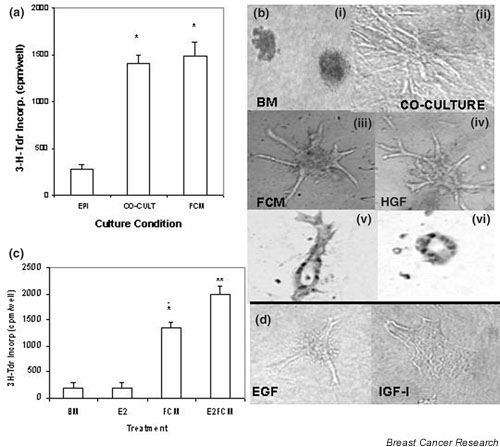
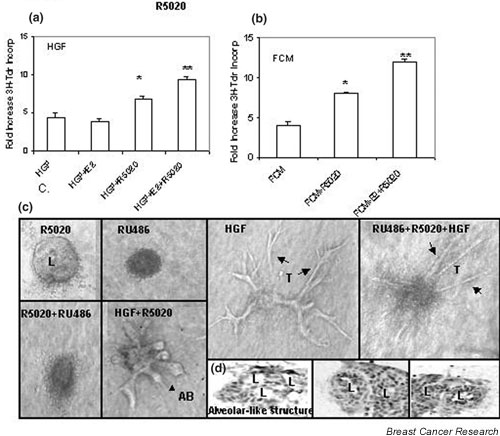
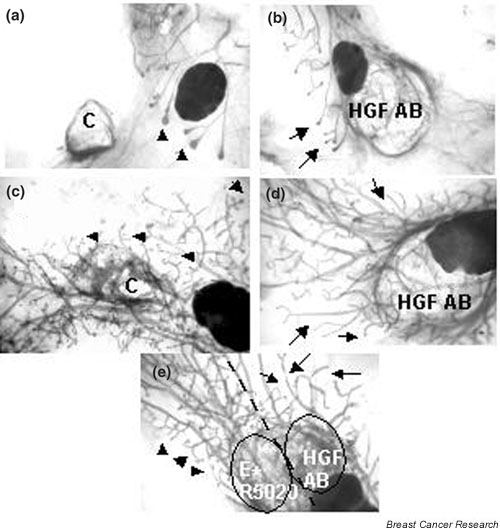
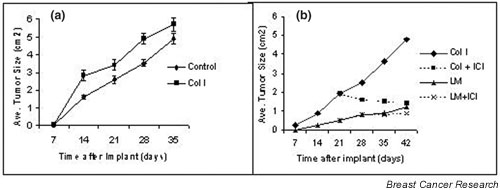
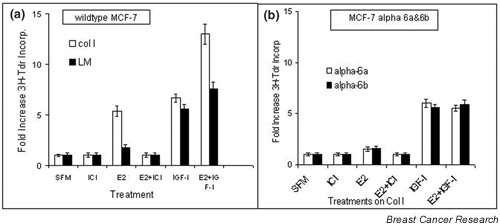
Mammary epithelial cells comprise the functional component of the normal gland and are the major target for carcinogenesis in mammary cancer. However, the stromal compartment of the normal gland and of tumors plays an important role in directing proliferative and functional changes in the epithelium. In vivo and in vitro studies of the murine mammary gland have provided insights into novel stroma-dependent mechanisms by which estrogen and progesterone action in the epithelium can be modulated by hepatocyte growth factor (HGF) and the extracellular matrix proteins, collagen type I, fibronectin and laminin. In vitro and in vivo studies of estrogen receptor positive, estrogen-responsive human breast cancer cells have also demonstrated that estrogen responsiveness of tumor cells can also be modulated by extracellular matrix proteins, collagen type I and laminin.
Figures
References
-
- Hofseth LJ, Raafat AM, Osuch JR, Slomski C. Effects of hormone replacement therapy with estrogen or estrogen plus progestagen on normal breast tissue in post-menopausal women. J Clin Endocrinol Metab. 1999;84:4559–4565. - PubMed
-
- Stadel BV. Hormone replacement therapy and risk of breast cancer. JAMA. 2002;287:2360–2361. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
