

Thioredoxin from Brugia malayi: defining a 16-kilodalton class of thioredoxins from nematodes
- PMID: 12819103
- PMCID: PMC162031
- DOI: 10.1128/IAI.71.7.4119-4126.2003
Thioredoxin from Brugia malayi: defining a 16-kilodalton class of thioredoxins from nematodes
Abstract
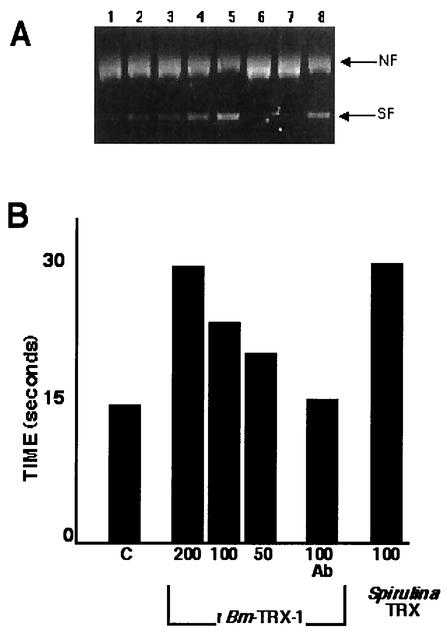
Thioredoxins are a family of small redox proteins that undergo NADPH-dependent reduction by thioredoxin reductase. This results in a supply of reducing equivalents that cells use in a wide variety of biological reactions, which include maintaining reduced forms of the enzymes important for protection against damage from high-energy oxygen radicals, the regulation of transcription factor activity, and the inhibition of apoptosis. Here we report on a new member of the thioredoxin family of proteins from the filarial nematode Brugia malayi, Bm-TRX-1, which defines a new subclass of 16-kDa thioredoxins that occur widely in nematodes, including Caenorhabditis elegans. In addition to being larger than the thioredoxins found in mammalian and bacterial species, the putative active site sequence of Bm-TRX-1, WCPPC, does not conform to the highly conserved WCGPC reported for thioredoxins from mammals to bacteria. Interestingly, an allelic form of Bm-TRX-1 was identified with an active site sequence WCPQC, which appears to be unique to the thioredoxins from filarial species. Bm-TRX-1 was between 98% and 35% identical to thioredoxins from other nematodes and approximately 20% identical to the thioredoxins from mammals and Escherichia coli. Bm-TRX-1 was constitutively transcribed throughout the B. malayi life cycle, and Bm-TRX protein was detectable in somatic extracts and excretory-secretory products from adults and microfilariae. Recombinant Bm-TRX-1 had thiodisulfide reductase activity, as measured by the reduction of insulin, and protected DNA from the nicking activity of oxygen radicals. Overexpression of Bm-TRX-1 in a human monocyte cell line negatively regulated tumor necrosis factor alpha-induced p38 mitogen-activated protein kinase activity, suggesting a possible role of the 16-kDa Bm-TRX-1 in immunomodulation.
Figures





Similar articles
-
Thioredoxin peroxidases from Brugia malayi.Mol Biochem Parasitol. 1998 Mar 15;91(2):207-20. doi: 10.1016/s0166-6851(97)00213-2. Mol Biochem Parasitol. 1998. PMID: 9566515
-
The role of a Brugia malayi p38 MAP kinase ortholog (Bm-MPK1) in parasite anti-oxidative stress responses.Mol Biochem Parasitol. 2011 Apr;176(2):90-7. doi: 10.1016/j.molbiopara.2010.12.008. Epub 2010 Dec 24. Mol Biochem Parasitol. 2011. PMID: 21185874
-
Biochemical and structural characterizations of thioredoxin reductase selenoproteins of the parasitic filarial nematodes Brugia malayi and Onchocerca volvulus.Redox Biol. 2022 May;51:102278. doi: 10.1016/j.redox.2022.102278. Epub 2022 Mar 4. Redox Biol. 2022. PMID: 35276442 Free PMC article.
-
Thioredoxin and related proteins in procaryotes.FEMS Microbiol Rev. 1988 Dec;4(4):271-97. doi: 10.1111/j.1574-6968.1988.tb02747.x. FEMS Microbiol Rev. 1988. PMID: 3152490 Review.
-
Properties and biological activities of thioredoxins.Annu Rev Pharmacol Toxicol. 2001;41:261-95. doi: 10.1146/annurev.pharmtox.41.1.261. Annu Rev Pharmacol Toxicol. 2001. PMID: 11264458 Review.
Cited by
-
Modeling analysis of GST (glutathione-S-transferases) from Wuchereria bancrofti and Brugia malayi.Bioinformation. 2005 Jun 2;1(1):25-7. doi: 10.6026/97320630001025. Bioinformation. 2005. PMID: 17597848 Free PMC article.
-
Stage- and gender-specific proteomic analysis of Brugia malayi excretory-secretory products.PLoS Negl Trop Dis. 2008;2(10):e326. doi: 10.1371/journal.pntd.0000326. Epub 2008 Oct 29. PLoS Negl Trop Dis. 2008. PMID: 18958170 Free PMC article.
-
Identification and characterization of a putative protein disulfide isomerase (HsPDI) as an alleged effector of Heterodera schachtii.Sci Rep. 2017 Oct 19;7(1):13536. doi: 10.1038/s41598-017-13418-9. Sci Rep. 2017. PMID: 29051538 Free PMC article.
-
Development of a Multiplex Bead Assay for the Detection of IgG Antibody Responses to Guinea Worm.Am J Trop Med Hyg. 2020 Dec;103(6):2294-2304. doi: 10.4269/ajtmh.20-0511. Epub 2020 Sep 3. Am J Trop Med Hyg. 2020. PMID: 32901602 Free PMC article.
-
The Secretome of Filarial Nematodes and Its Role in Host-Parasite Interactions and Pathogenicity in Onchocerciasis-Associated Epilepsy.Front Cell Infect Microbiol. 2021 Apr 28;11:662766. doi: 10.3389/fcimb.2021.662766. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33996633 Free PMC article. Review.
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Arner, E. S., and A. Holmgren. 2000. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 267:6102-6109. - PubMed
-
- Berggren, M., A. Gallegos, J. R. Gasdaska, P. Y. Gasdaska, J. Warneke, and G. Powis. 1996. Thioredoxin and thioredoxin reductase gene expression in human tumors and cell lines, and the effects of serum stimulation and hypoxia. Anticancer Res. 16:3459-3466. - PubMed
-
- Bertini, R., O. M. Howard, H. F. Dong, J. J. Oppenheim, C. Bizzarri, R. Sergi, G. Caselli, S. Pagliei, B. Romines, J. A. Wilshire, M. Mengozzi, H. Nakamura, J. Yodoi, K. Pekkari, R. Gurunath, A. Holmgren, L. A. Herzenberg, and P. Ghezzi. 1999. Thioredoxin, a redox enzyme released in infection and inflammation, is a unique chemoattractant for neutrophils, monocytes, and T cells. J. Exp. Med. 189:1783-1789. - PMC - PubMed
-
- Blaxter, M., and M. Aslett. 1997. Internet resources for the parasite genome projects. Trends Genet. 13:40-41. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
