

Analysis of Tat transactivation of human immunodeficiency virus transcription in vitro
- PMID: 1282057
- PMCID: PMC6057369
Analysis of Tat transactivation of human immunodeficiency virus transcription in vitro
Abstract
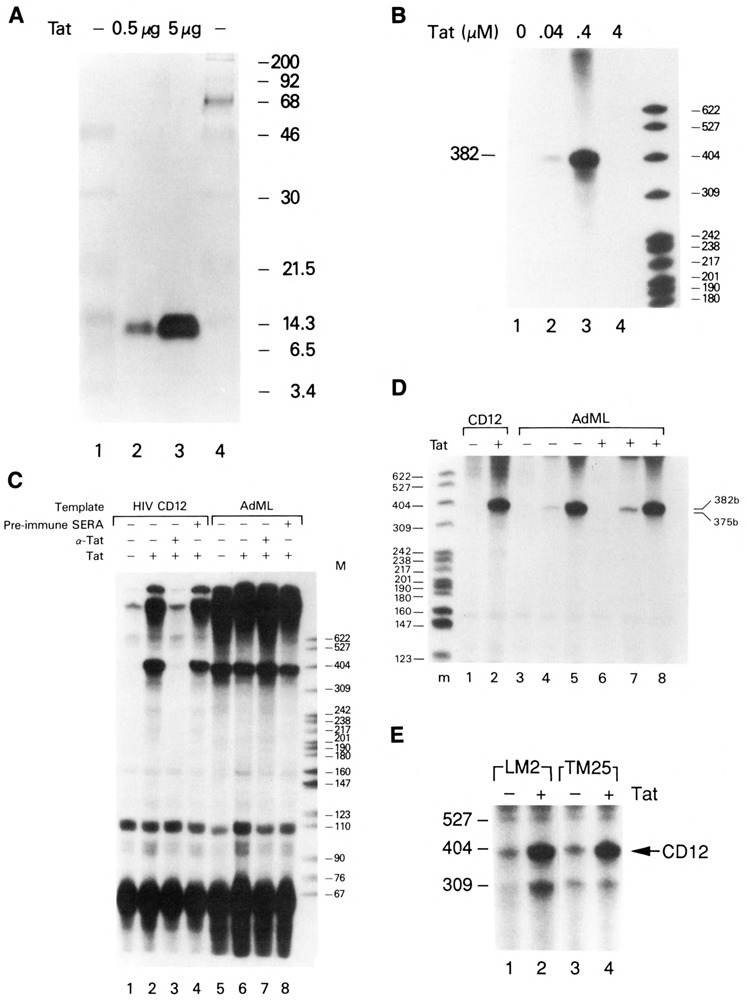
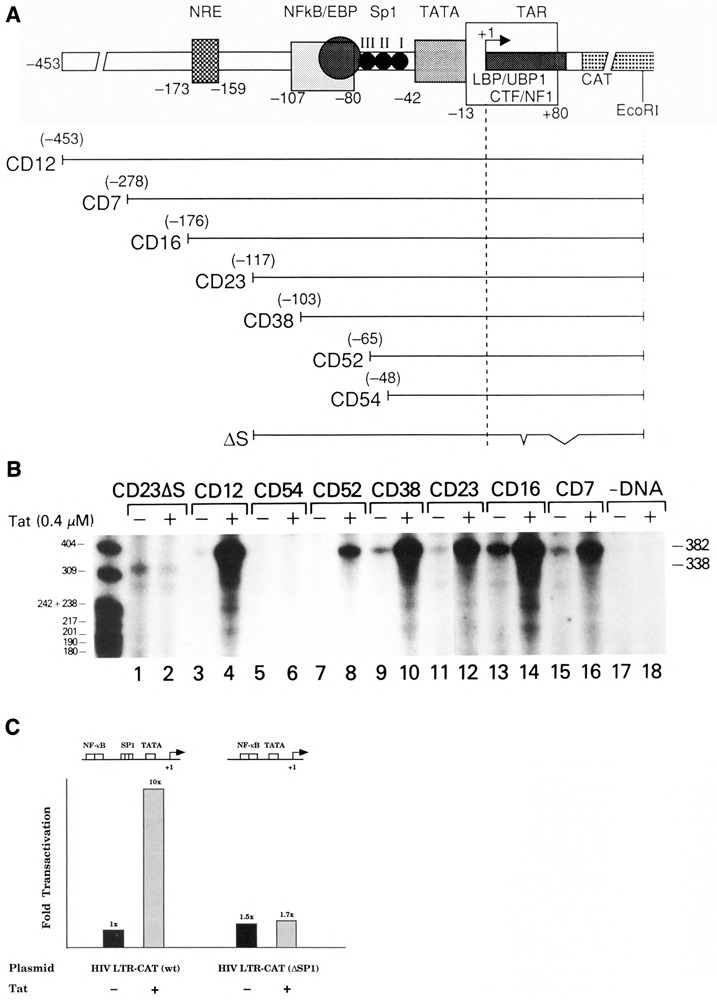
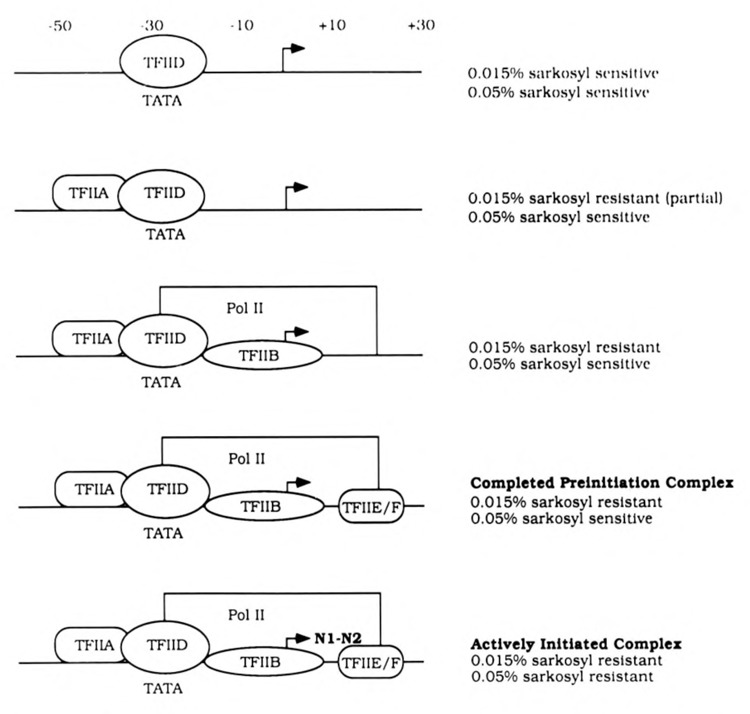
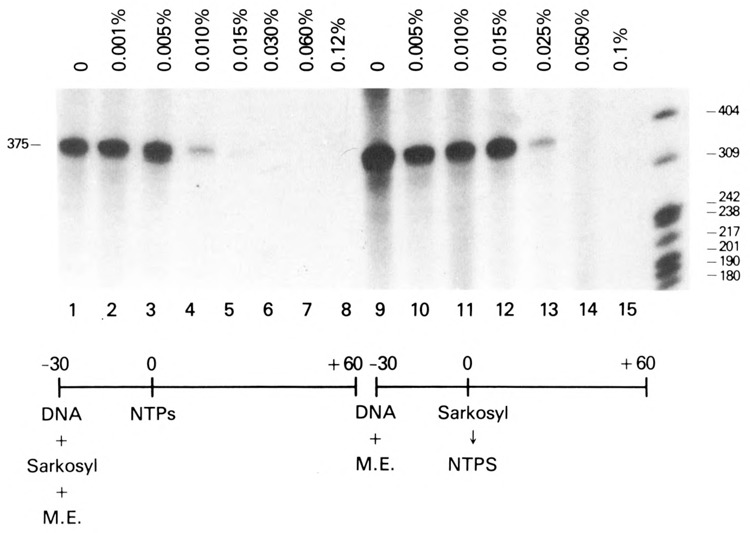
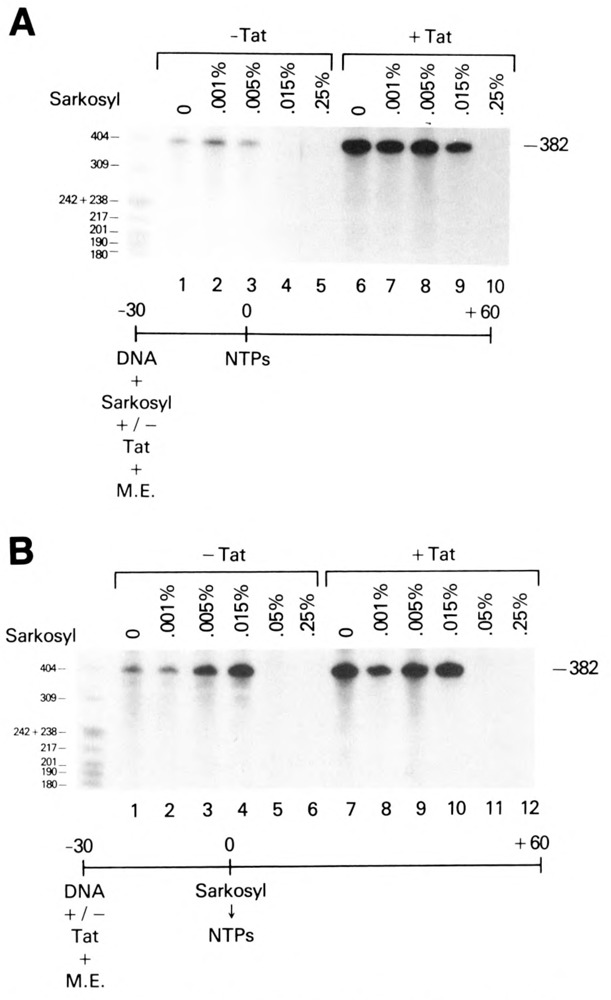
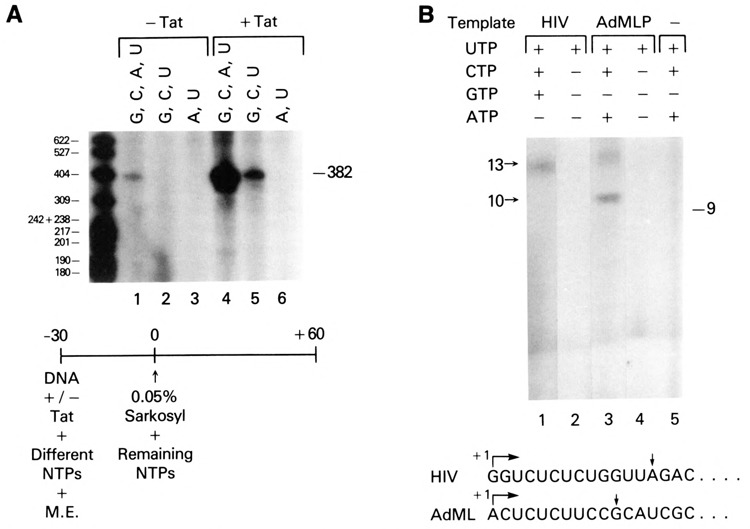
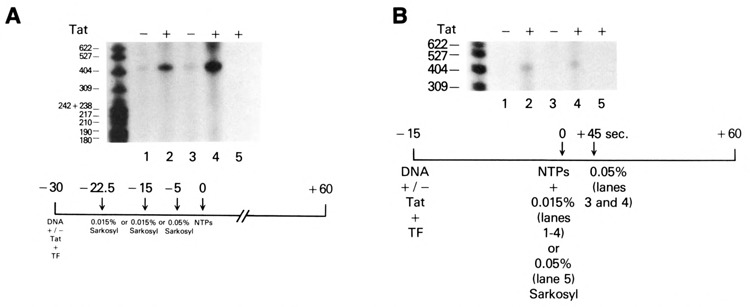
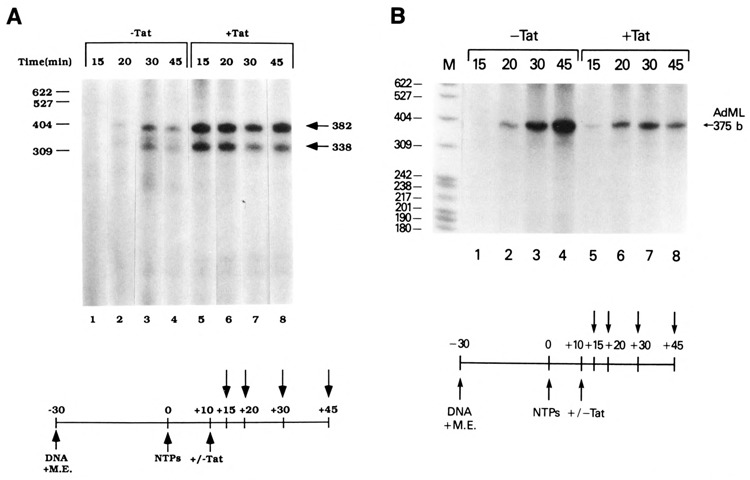
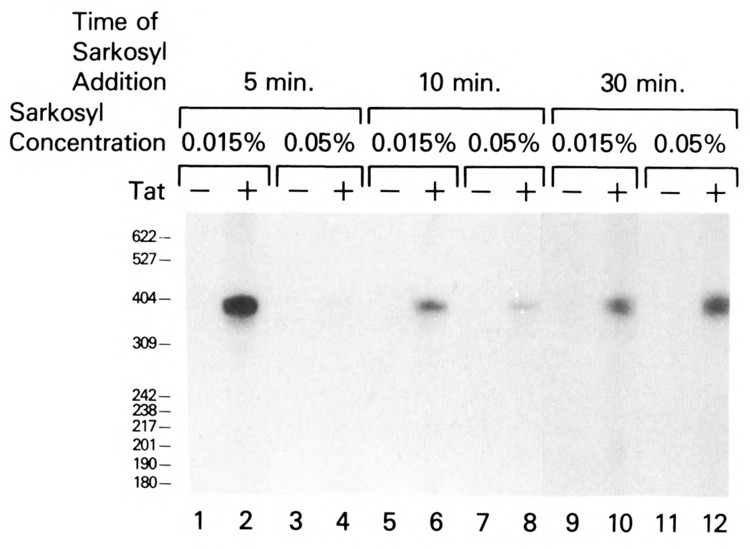
The HIV Tat protein is a potent transactivator of HIV transcription, increasing both RNA initiation and elongation. We now demonstrate that purified, full-length 86 amino acid Tat protein specifically transactivates the HIV LTR in vitro to a high level (25- to 60-fold). Tat transactivation was specifically blocked by anti-Tat serum, but not preimmune serum. Tat did not transactivate transcription from the control adenovirus major late promoter (AdMLP). HIV transcription was blocked at various functional steps during initiation and elongation complex formation. Similar to the control AdMLP, HIV basal initiation complex assembly was sensitive to the addition of 0.015% sarkosyl prior to the addition of nucleoside triphosphates. Resistance to 0.05% sarkosyl required the addition of G, C, and U, which constitute the first 13 bases of the HIV RNA transcript. The addition of Tat to the in vitro transcription relieved the 0.015% sarkosyl block. These Tat-induced complexes were sensitive to 0.05% sarkosyl, suggesting that transcriptional initiation had not occurred. Consistent with this hypothesis, the addition of G, C, and U to the Tat-induced transcription complexes allowed the rapid conversion to transcription initiation complexes. Tat also facilitated the formation of 0.015% sarkosyl-resistant complexes in a reconstituted transcription system containing partially purified transcription factors and polymerase II. Following the formation of stable initiation complexes, Tat increased the rate and efficiency of transcription elongation on the HIV but not the AdML template. Kinetic analysis of Tat transactivation suggests that approximately 30% of the Tat initiation complexes are converted to elongation complexes. We conclude that Tat, in addition to its demonstrated role in RNA elongation, facilitates transcription initiation in vitro.
Figures
References
-
- Abmayr S. M., Workman J. L., and Roeder R. G. (1988), Genes Dev 2, 542–553. - PubMed
-
- Arya S. K., Guo C., Josephs S. F., and Wong-Staal F. (1985), Science 229, 69–73. - PubMed
-
- Barre-Sinoussi T., Chermann J. C., Reye F., Nugeure M. T., Charmaret S., Gruest J., Dauquet C., Aixer-Blin C., Vezinet-Brun F., Rouzious C., Rozenbaum W., and Montagnier L. (1983), Science 220, 868–871. - PubMed
-
- Berkhout B., Gatignol A., Rabson A. B., and Jeang K.-T. (1990), Cell 62, 757–767. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources