

Dynamic instability of microtubules is regulated by force
- PMID: 12821641
- PMCID: PMC2173003
- DOI: 10.1083/jcb.200301147
Dynamic instability of microtubules is regulated by force
Abstract
Microtubules are long filamentous protein structures that randomly alternate between periods of elongation and shortening in a process termed dynamic instability. The average time a microtubule spends in an elongation phase, known as the catastrophe time, is regulated by the biochemical machinery of the cell throughout the cell cycle. In this light, observed changes in the catastrophe time near cellular boundaries (Brunner, D., and P. Nurse. 2000. Cell. 102:695-704; Komarova, Y.A., I.A. Vorobjev, and G.G. Borisy. 2002. J. Cell Sci. 115:3527-3539) may be attributed to regulatory effects of localized proteins. Here, we argue that the pushing force generated by a microtubule when growing against a cellular object may itself provide a regulatory mechanism of the catastrophe time. We observed an up to 20-fold, force-dependent decrease in the catastrophe time when microtubules grown from purified tubulin were polymerizing against microfabricated barriers. Comparison with catastrophe times for microtubules growing freely at different tubulin concentrations leads us to conclude that force reduces the catastrophe time only by limiting the rate of tubulin addition.
Figures

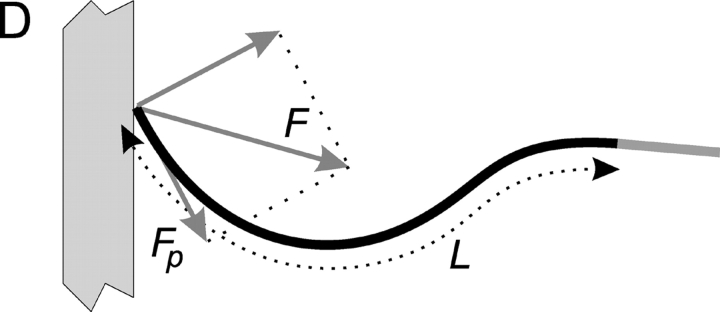
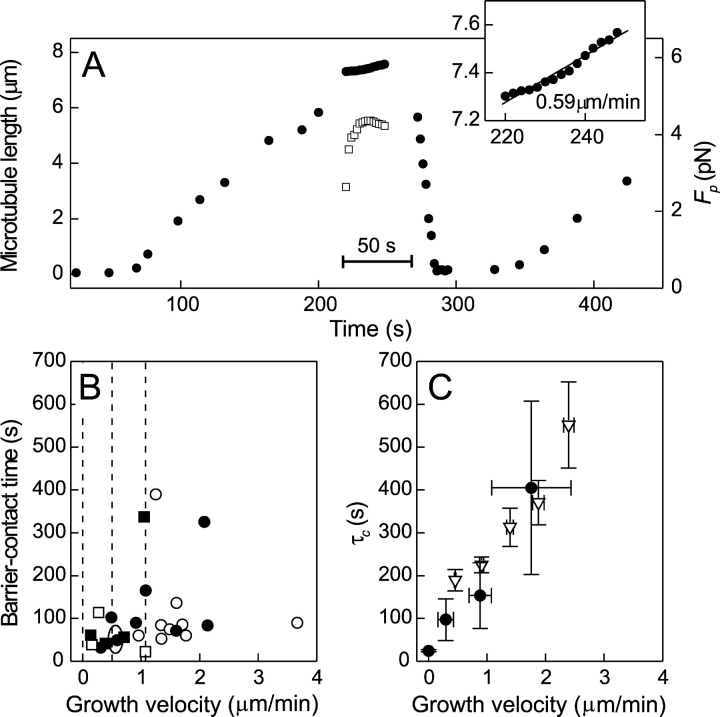
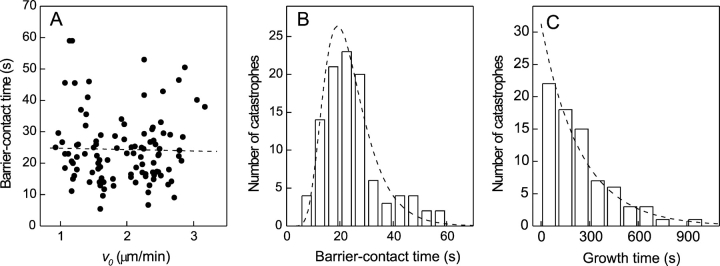
References
-
- Brunner, D., and P. Nurse. 2000. CLIP170-like tip1p spatially organizes microtubular dynamics in fission yeast. Cell. 102:695–704. - PubMed
-
- Desai, A., and T.J. Mitchison. 1997. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 13:83–117. - PubMed
-
- Dogterom, M., and B. Yurke. 1997. Measurement of the force-velocity relation for growing microtubules. Science. 278:856–860. - PubMed
-
- Dogterom, M., M.E. Janson, C. Faivre-Moskalenko, A. Van der Horst, J.W.J. Kerssemakers, C. Tanase, and B.M. Mulder. 2002. Force generation by polymerizing microtubules. Appl. Physics A-Mater. Sci. Process. 75:331–336.
-
- Doorn, G.S.v., C. Tanase, B.M. Mulder, and M. Dogterom. 2000. On the stall force for growing microtubules. Eur. Biophys. J. Biophys. Lett. 29:2–6. - PubMed
