

Low density lipoprotein receptor-related protein is a calreticulin coreceptor that signals focal adhesion disassembly
- PMID: 12821648
- PMCID: PMC2172996
- DOI: 10.1083/jcb.200302069
Low density lipoprotein receptor-related protein is a calreticulin coreceptor that signals focal adhesion disassembly
Erratum in
- J Cell Biol. 2003 Aug 4;162(3):521
Abstract
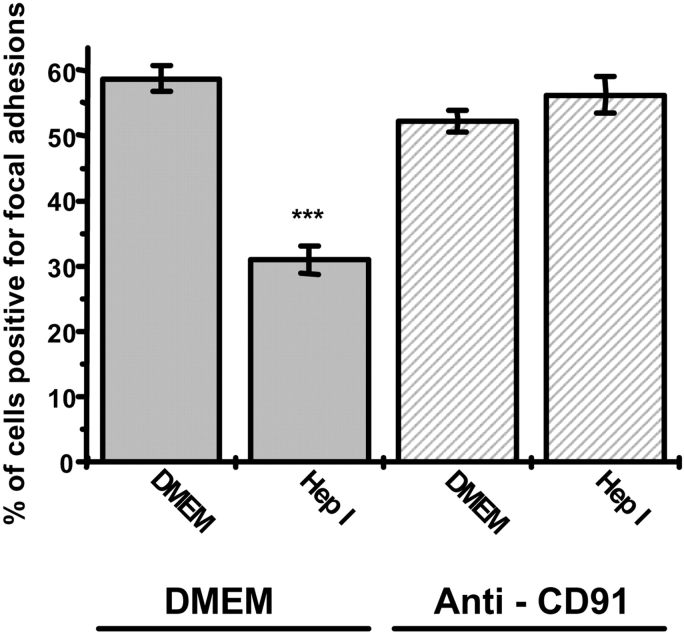
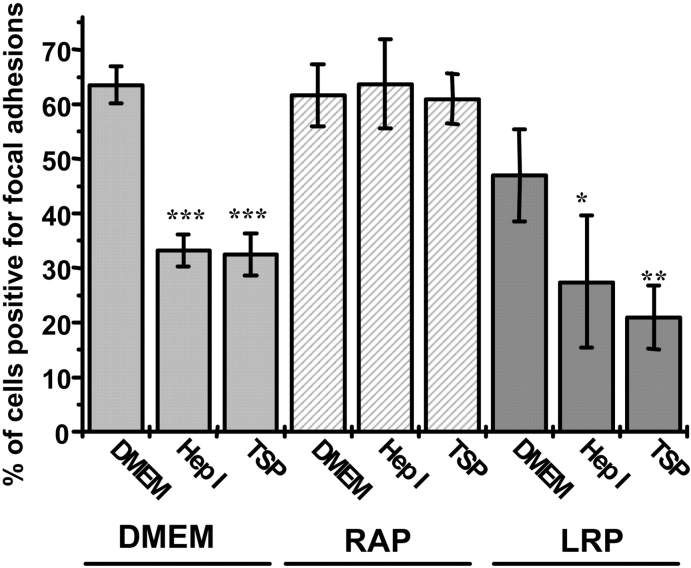
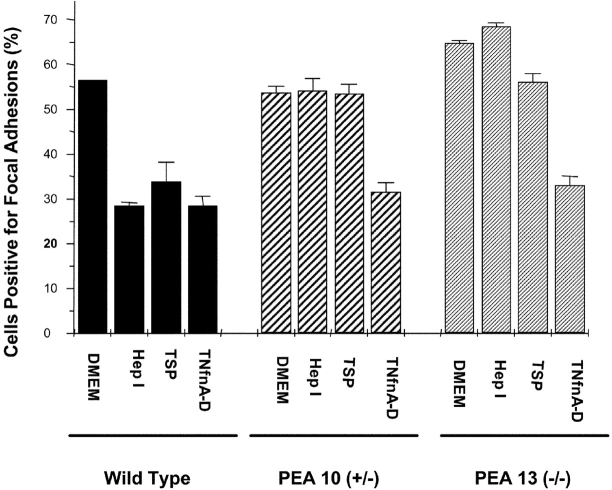
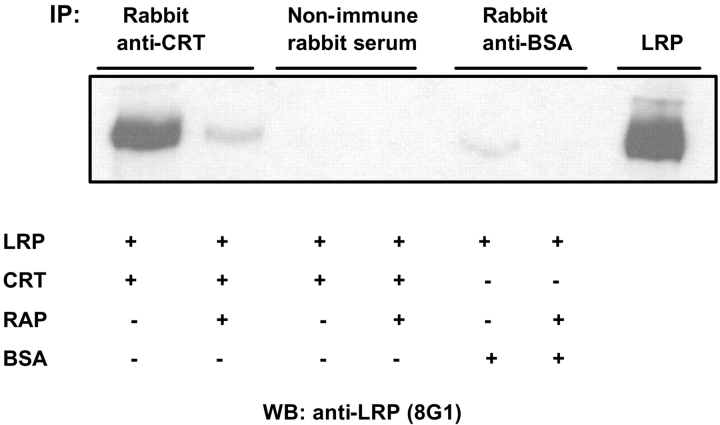
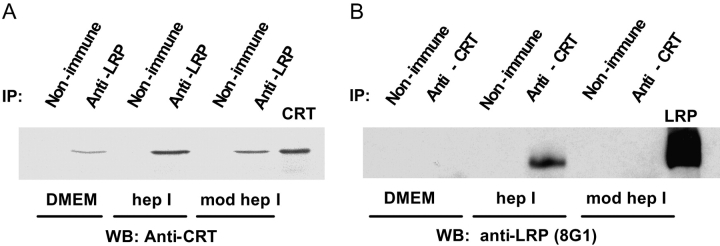
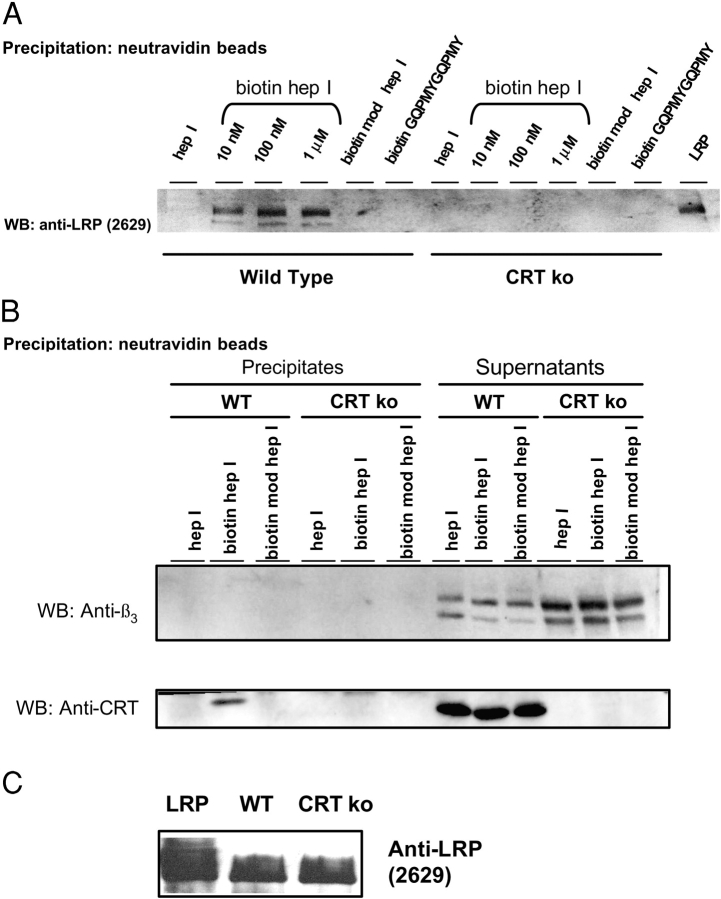
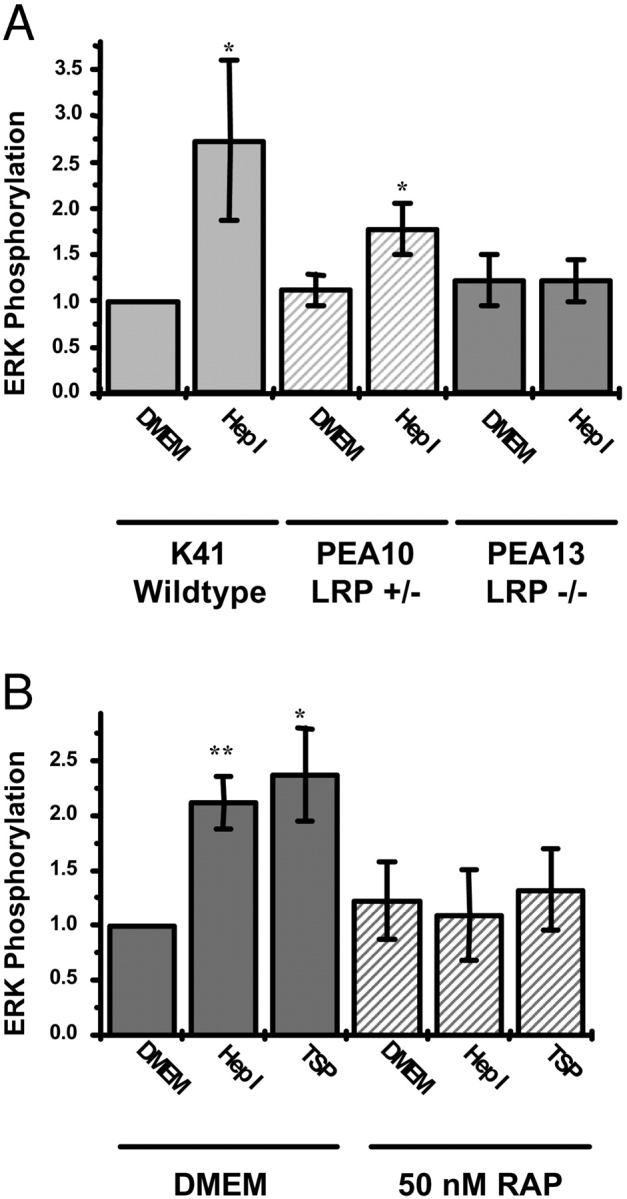
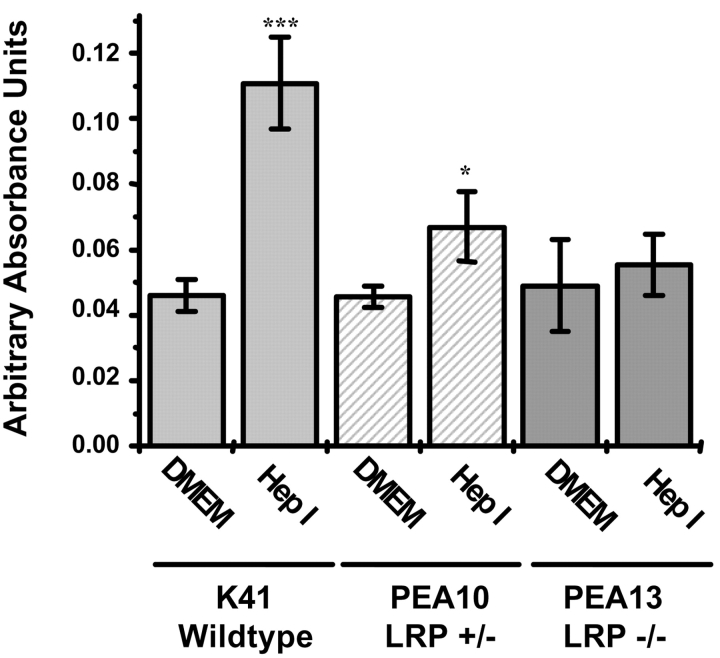
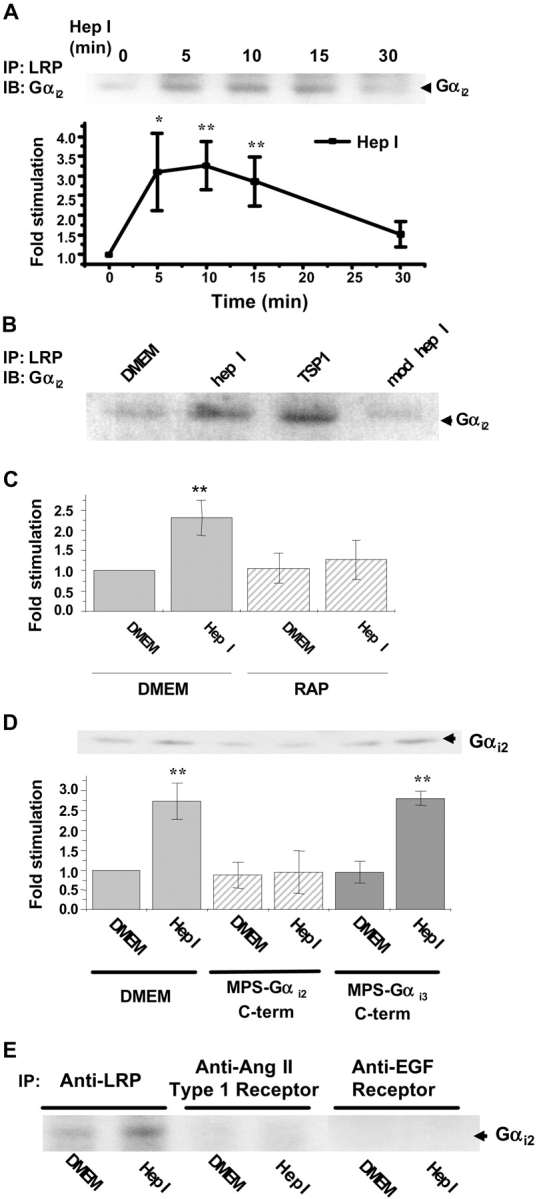
Thrombospondin (TSP) signals focal adhesion disassembly (the intermediate adhesive state) through interactions with cell surface calreticulin (CRT). TSP or a peptide (hep I) of the active site induces focal adhesion disassembly through binding to CRT, which activates phosphoinositide 3-kinase (PI3K) and extracellular signal-related kinase (ERK) through Galphai2 proteins. Because CRT is not a transmembrane protein, it is likely that CRT signals as part of a coreceptor complex. We now show that low density lipoprotein receptor-related protein (LRP) mediates focal adhesion disassembly initiated by TSP binding to CRT. LRP antagonists (antibodies, receptor-associated protein) block hep I/TSP-induced focal adhesion disassembly. LRP is necessary for TSP/hep I signaling because TSP/hep I is unable to stimulate focal adhesion disassembly or ERK or PI3K signaling in fibroblasts deficient in LRP. LRP is important in TSP-CRT signaling, as shown by the ability of hep I to stimulate association of Galphai2 with LRP. The isolated proteins LRP and CRT interact, and LRP and CRT are associated with hep I in molecular complexes extracted from cells. These data establish a mechanism of cell surface CRT signaling through its coreceptor, LRP, and suggest a novel function for LRP in regulating cell adhesion.
Figures
References
-
- Arosa, F.A., O. de Jesus, G. Porto, A.M. Carmo, and M. de Sousa. 1999. Calreticulin is expressed on the cell surface of activated human peripheral blood T lymphocytes in association with major histocompatibility complex class I molecules. J. Biol. Chem. 274:16917–16922. - PubMed
-
- Ashcom, J.D., S.E. Tiller, K. Dickerson, J.L. Cravens, W.S. Argraves, and D.K. Strickland. 1990. The human alpha 2-macroglobulin receptor: identification of a 420-kD cell surface glycoprotein specific for the activated conformation of alpha 2-macroglobulin. J. Cell Biol. 110:1041–1048. - PMC - PubMed
-
- Aukhil, I., P. Joshi, Y. Yan, and H.P. Erickson. 1993. Cell- and heparin-binding domains of the hexabrachion arm identified by tenascin expression proteins. J. Biol. Chem. 268:2542–2553. - PubMed
-
- Baksh, S., and M. Michalak. 1991. Expression of calreticulin in Escherichia coli and identification of its Ca2+ binding domains. J. Biol. Chem. 266:21458–21465. - PubMed
-
- Barnes, H., B. Larsen, M. Tyers, and P. van Der Geer. 2001. Tyrosine-phosphorylated low density lipoprotein receptor-related protein 1 (LRP1) associates with the adapter protein SHC in SRC-transformed cells. J. Biol. Chem. 276:19119–19125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
