

Comparison of calculation and experiment implicates significant electrostatic contributions to the binding stability of barnase and barstar
- PMID: 12829463
- PMCID: PMC1303064
- DOI: 10.1016/S0006-3495(03)74453-1
Comparison of calculation and experiment implicates significant electrostatic contributions to the binding stability of barnase and barstar
Abstract
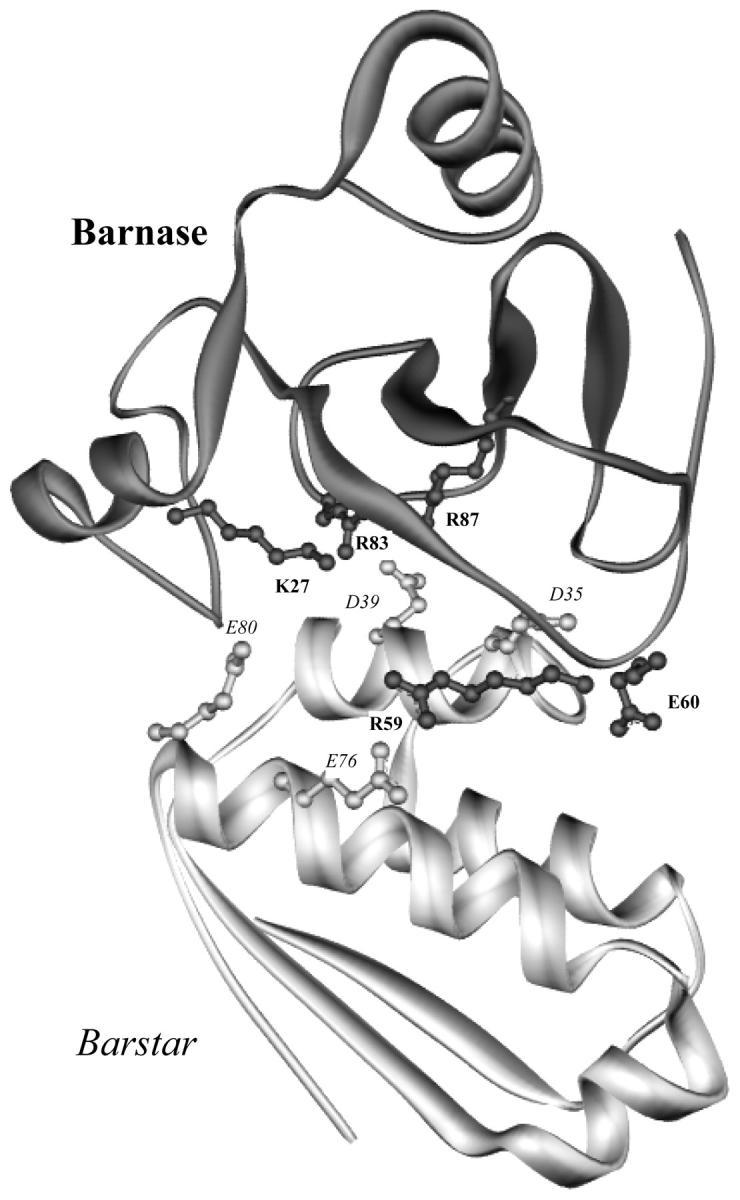
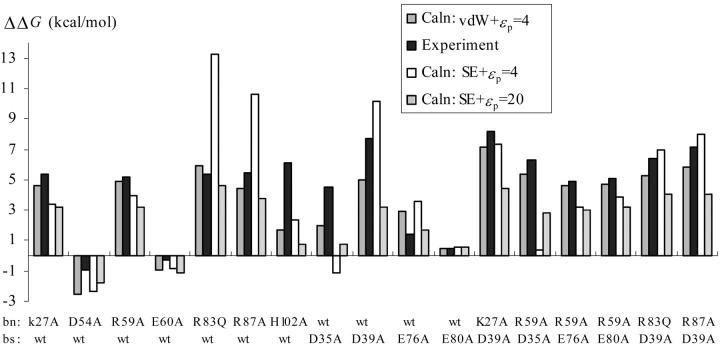
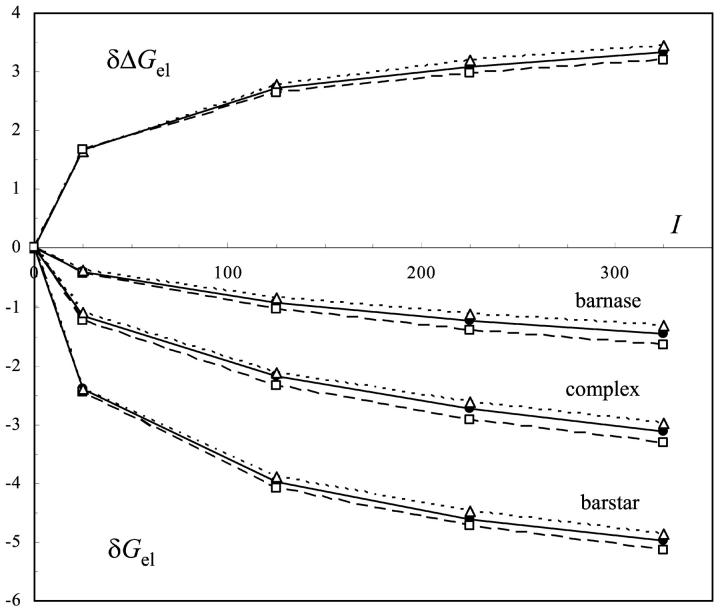
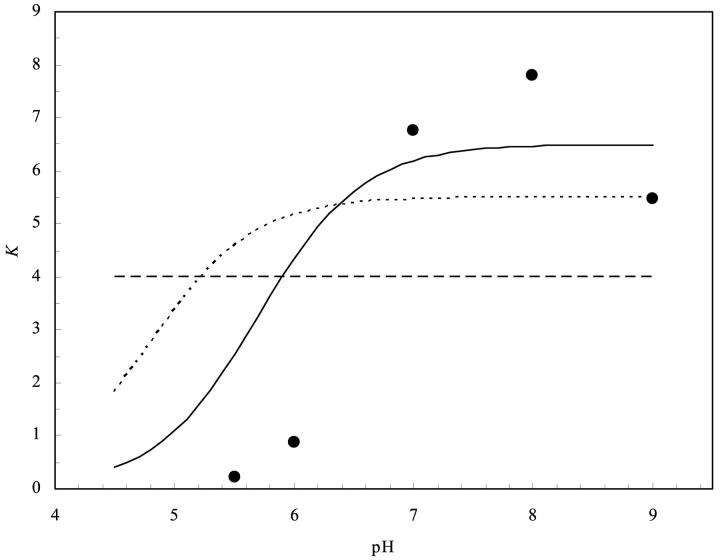
The contributions of electrostatic interactions to the binding stability of barnase and barstar were studied by the Poisson-Boltzmann model with three different protocols: a), the dielectric boundary specified as the van der Waals (vdW) surface of the protein along with a protein dielectric constant (epsilon (p)) of 4; b), the dielectric boundary specified as the molecular (i.e., solvent-exclusion (SE)) surface along with epsilon (p) = 4; and c), "SE + epsilon (p) = 20." The "vdW + epsilon (p) = 4" and "SE + epsilon (p) = 20" protocols predicted an overall electrostatic stabilization whereas the "SE + epsilon (p) = 4" protocol predicted an overall electrostatic destabilization. The "vdW + epsilon (p) = 4" protocol was most consistent with experiment. It quantitatively reproduced the observed effects of 17 mutations neutralizing charged residues lining the binding interface and the measured coupling energies of six charge pairs across the interface and reasonably rationalized the experimental ionic strength and pH dependences of the binding constant. In contrast, the "SE + epsilon (p) = 4" protocol predicted significantly larger coupling energies of charge pairs whereas the "SE + epsilon (p) = 20" protocol did not predict any pH dependence. This study calls for further scrutiny of the different Poisson-Boltzmann protocols and demonstrates potential danger in drawing conclusions on electrostatic contributions based on a particular calculation protocol.
Figures


References
-
- Albeck, S., R. Unger, and G. Schreiber. 2000. Evaluation of direct and cooperative contributions towards the strength of buried hydrogen bonds and salt bridges. J. Mol. Biol. 298:503–520. - PubMed
-
- Alexov, E. 2003. Role of the protein side-chain fluctuations on the strength of pair-wise electrostatic interactions: comparing experimental with computed pKas. Proteins. 50:94–103. - PubMed
-
- Antosiewicz, J., J. A. McCammon, and M. K. Gilson. 1994. Prediction of pH-dependent properties of proteins. J. Mol. Biol. 238:415–436. - PubMed
-
- Antosiewicz, J., J. A. McCammon, and M. K. Gilson. 1996. The determinants of pKas in proteins. Biochemistry. 35:7819–7833. - PubMed
-
- Buckle, A. M., and A. R. Fersht. 1994. Subsite binding in an RNase: structure of a barnase-tetranucleotide complex at 1.76-A resolution. Biochemistry. 33:1644–1653. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
