

C/EBPbeta regulation in lipopolysaccharide-stimulated macrophages
- PMID: 12832471
- PMCID: PMC162211
- DOI: 10.1128/MCB.23.14.4841-4858.2003
C/EBPbeta regulation in lipopolysaccharide-stimulated macrophages
Abstract
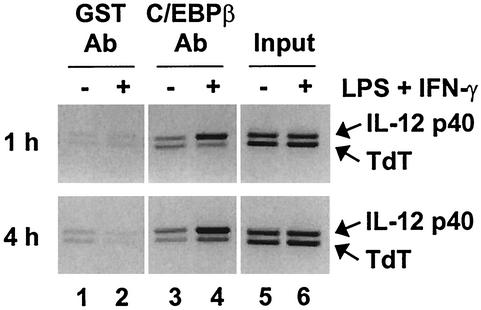
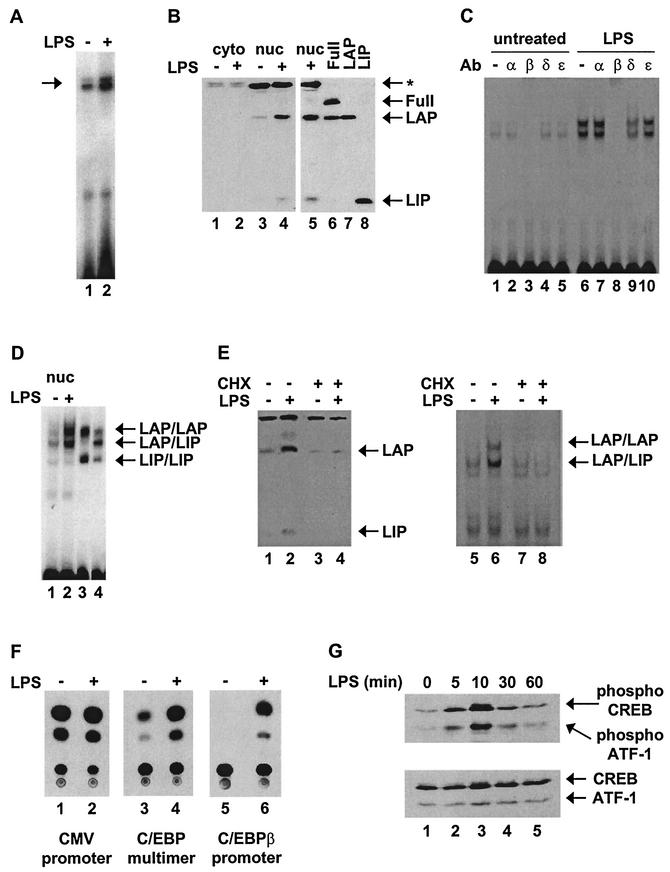
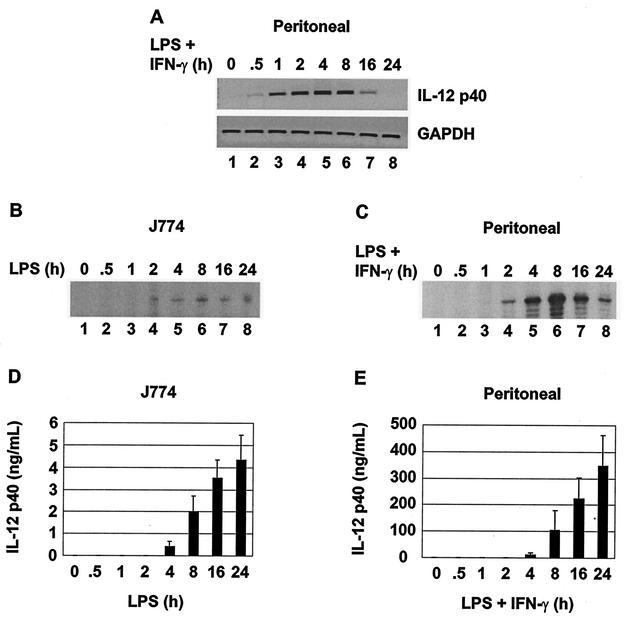
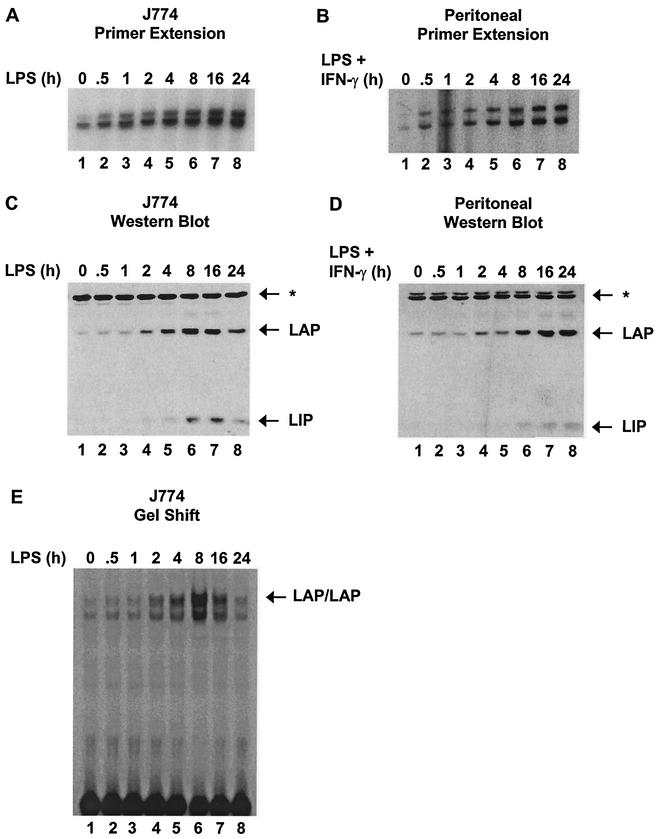
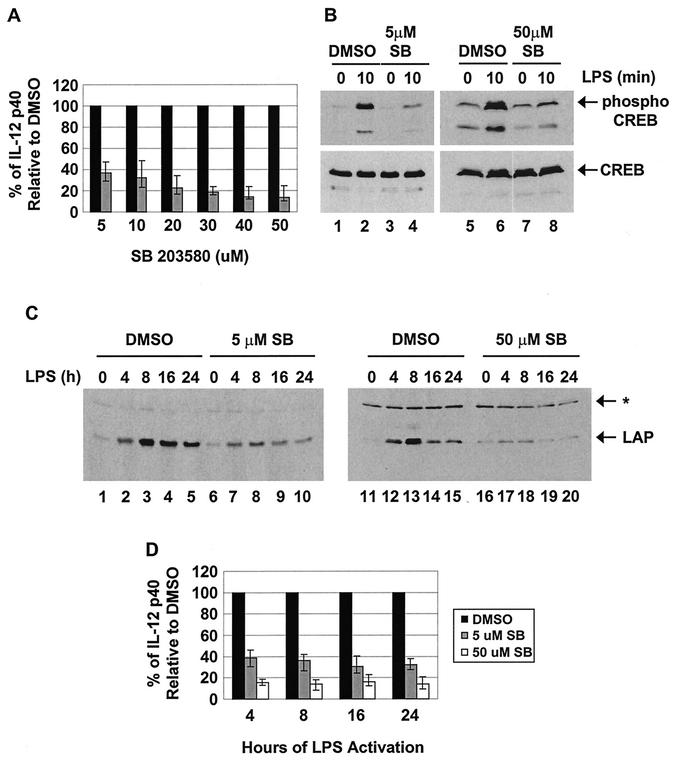
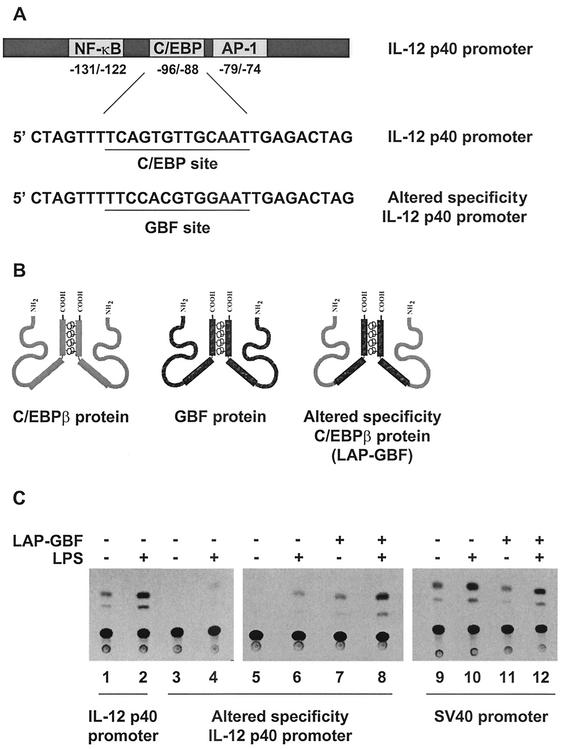
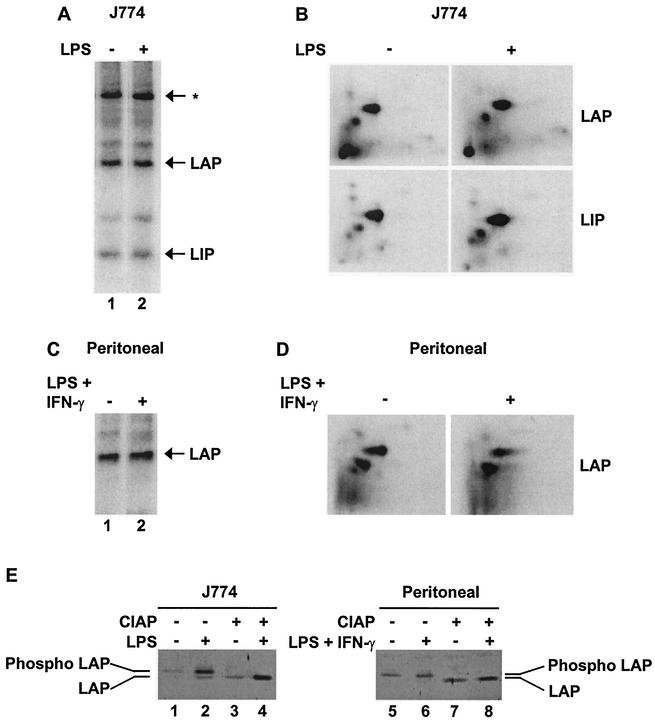
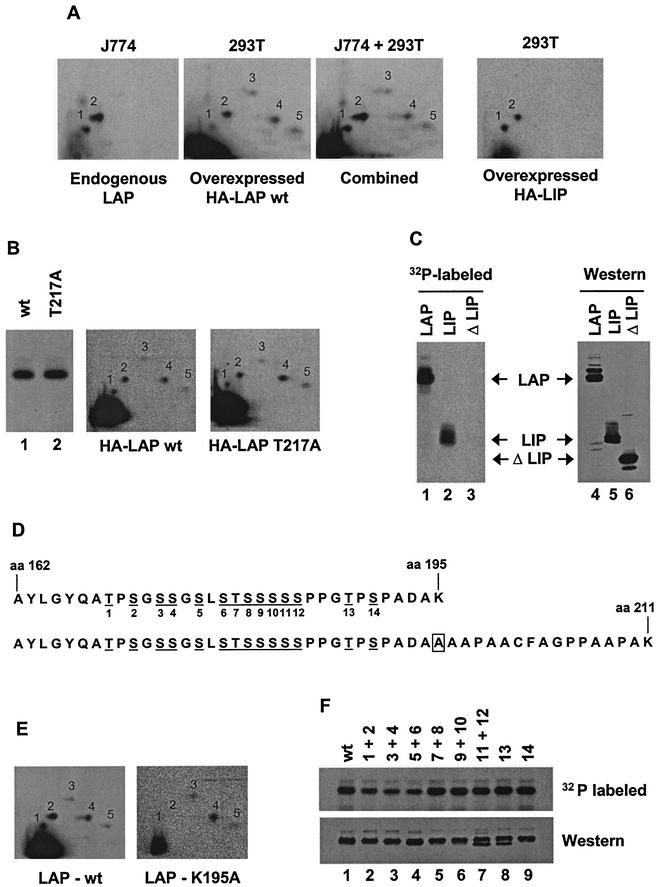
C/EBP family members contribute to the induction of the interleukin-12 p40 gene and the genes encoding several other mediators of inflammation. Here, we show by chromatin immunoprecipitation that C/EBPbeta binds the p40 promoter following lipopolysaccharide stimulation of peritoneal macrophages. However, three modes of C/EBPbeta regulation reported in other cell types were not detected, including alternative translation initiation, nuclear translocation, and increased DNA binding following posttranslational modification. In contrast, C/EBPbeta concentrations greatly increased following stimulation via MAP kinase-dependent induction of C/EBPbeta gene transcription. Increased C/EBPbeta concentrations were unimportant for p40 induction, however, as transcription of the p40 gene initiated before C/EBPbeta concentrations increased. Furthermore, disruption of C/EBPbeta upregulation by a MAP kinase inhibitor only slightly diminished p40 induction. Phosphopeptide mapping revealed that endogenous C/EBPbeta in macrophages is phosphorylated on only a single tryptic peptide containing 14 potential phosphoacceptors. This peptide was constitutively phosphorylated in primary and transformed macrophages, in contrast to its inducible phosphorylation in other cell types in response to Ras and growth hormone signaling. Altered-specificity experiments supported the hypothesis that C/EBPbeta activity in macrophages does not require an inducible posttranslational modification. These findings suggest that, although C/EBPbeta contributes to the induction of numerous proinflammatory genes, it is fully active in unstimulated macrophages and poised to stimulate transcription in conjunction with other factors whose activities are induced.
Figures
References
-
- Agre, P., P. F. Johnson, and S. L. McKnight. 1989. Cognate DNA binding specificity retained after leucine zipper exchange between GCN4 and C/EBP. Science 246:922-926. - PubMed
-
- Alam, T., M. R. An, and J. Papaconstantinou. 1992. Differential expression of three C/EBP isoforms in multiple tissues during the acute phase response. J. Biol. Chem. 267:5021-5024. - PubMed
-
- An, M. R., C.-C. Hsieh, P. D. Reisner, J. P. Rabek, S. G. Scott, D. T. Kuninger, and J. Papaconstantinou. 1996. Evidence for posttranslational regulation of C/EBPα and C/EBPβ isoform expression during the lipopolysaccharide-mediated acute-phase response. Mol. Cell. Biol. 16:2295-2306. - PMC - PubMed
-
- Berrier, A., G. Siu, and K. Calame. 1998. Transcription of a minimal promoter from the NF-IL6 gene is regulated by CREB/ATF and SP1 proteins in U937 promonocytic cells. J. Immunol. 161:2267-2275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources