

Self-organizing neural integrator predicts interval times through climbing activity
- PMID: 12832560
- PMCID: PMC6741165
- DOI: 10.1523/JNEUROSCI.23-12-05342.2003
Self-organizing neural integrator predicts interval times through climbing activity
Abstract
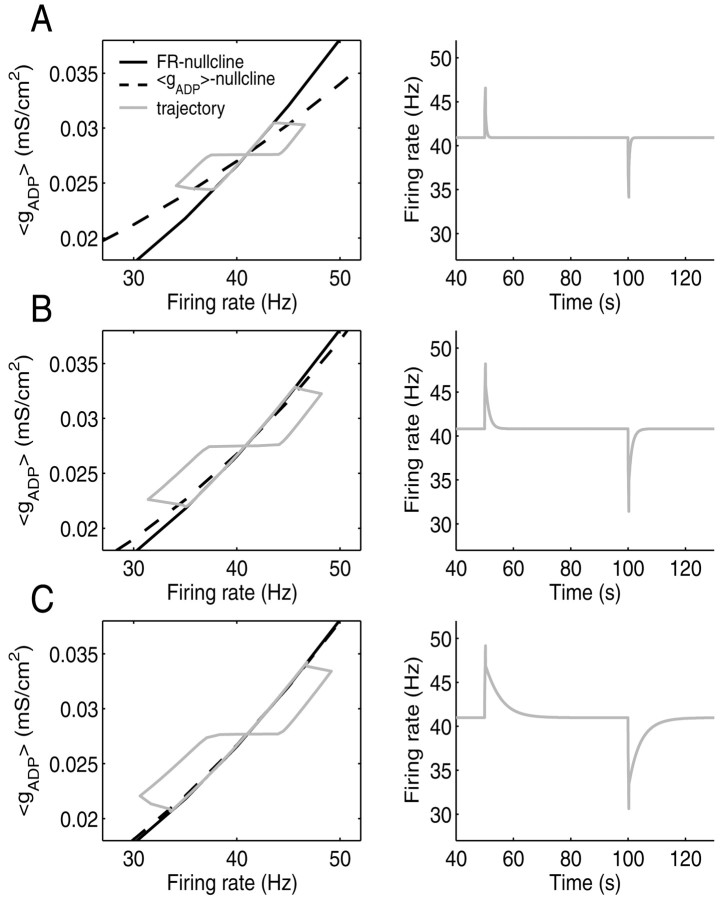
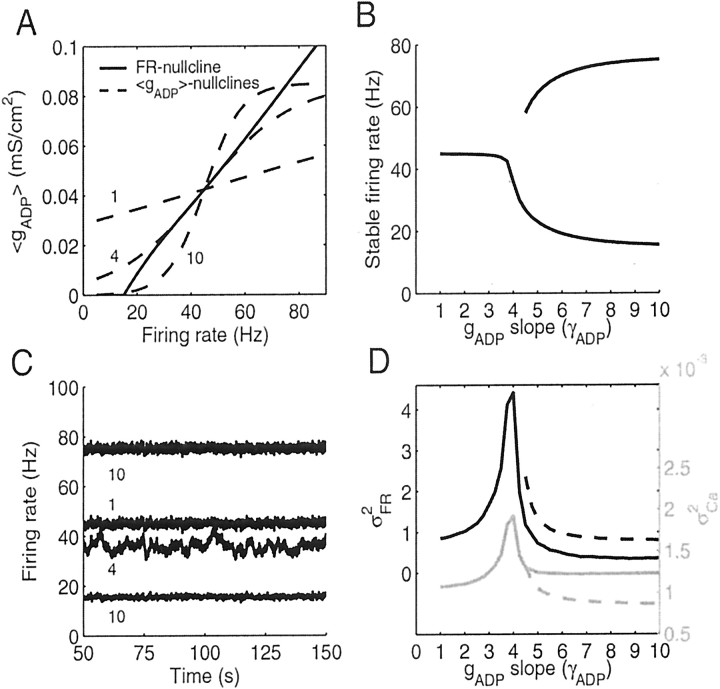
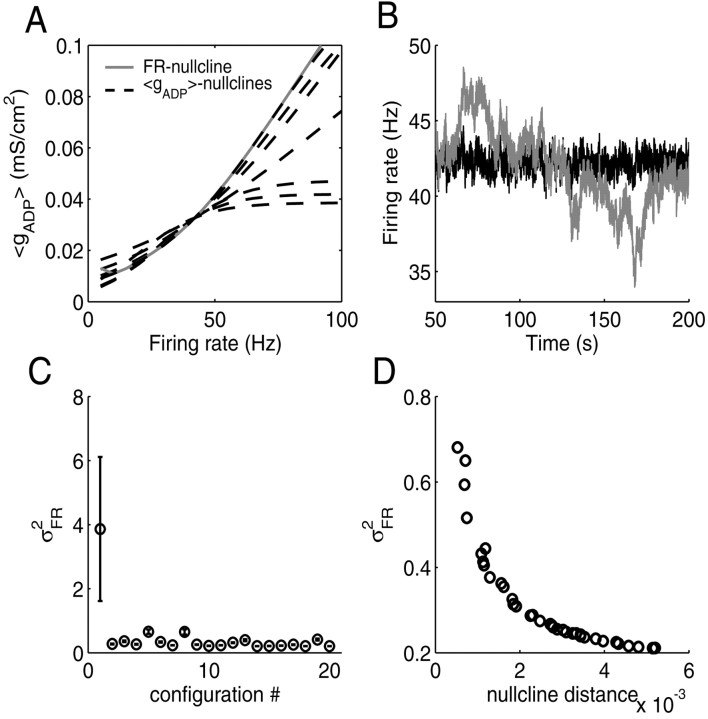
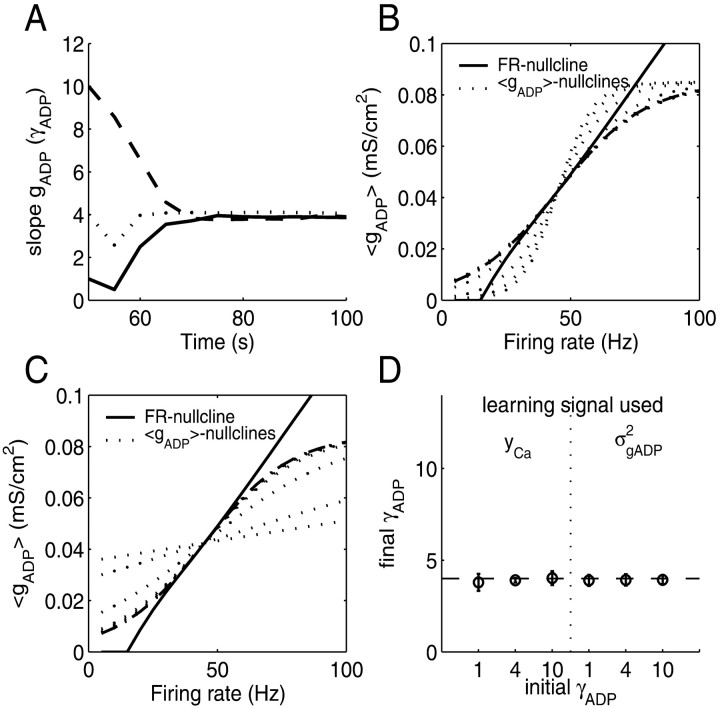
Mammals can reliably predict the time of occurrence of an expected event after a predictive stimulus. Climbing activity is a prominent profile of neural activity observed in prefrontal cortex and other brain areas that is related to the anticipation of forthcoming events. Climbing activity might span intervals from hundreds of milliseconds to tens of seconds and has a number of properties that make it a plausible candidate for representing interval time. A biophysical model is presented that produces climbing, temporal integrator-like activity with variable slopes as observed empirically, through a single-cell positive feedback loop between firing rate, spike-driven Ca2+ influx, and Ca2+-activated inward currents. It is shown that the fine adjustment of this feedback loop might emerge in a self-organizing manner if the cell can use the variance in intracellular Ca2+ fluctuations as a learning signal. This self-organizing process is based on the present observation that the variance of the intracellular Ca2+ concentration and the variance of the neural firing rate and of activity-dependent conductances reach a maximum as the biophysical parameters of a cell approach a configuration required for temporal integration. Thus, specific mechanisms are proposed for (1) how neurons might represent interval times of variable length and (2) how neurons could acquire the biophysical properties that enable them to work as timers.
Figures
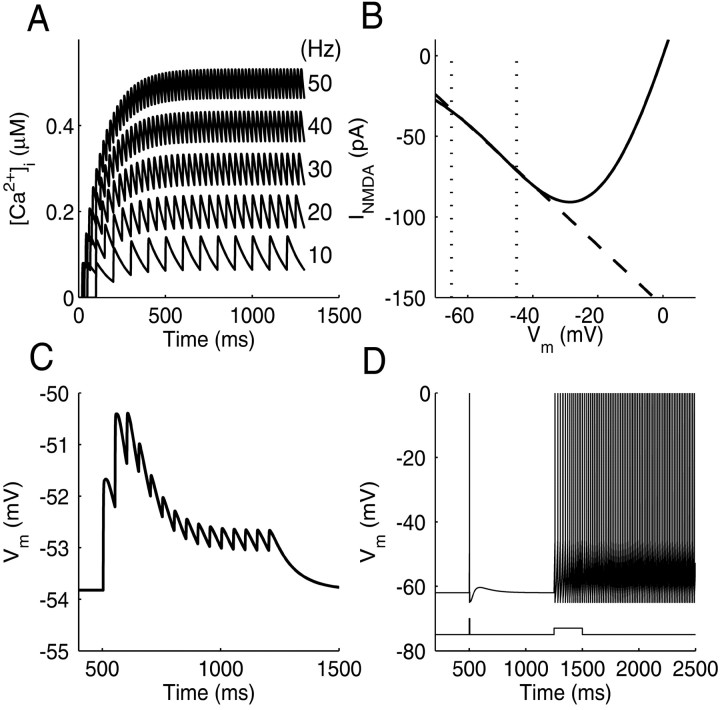
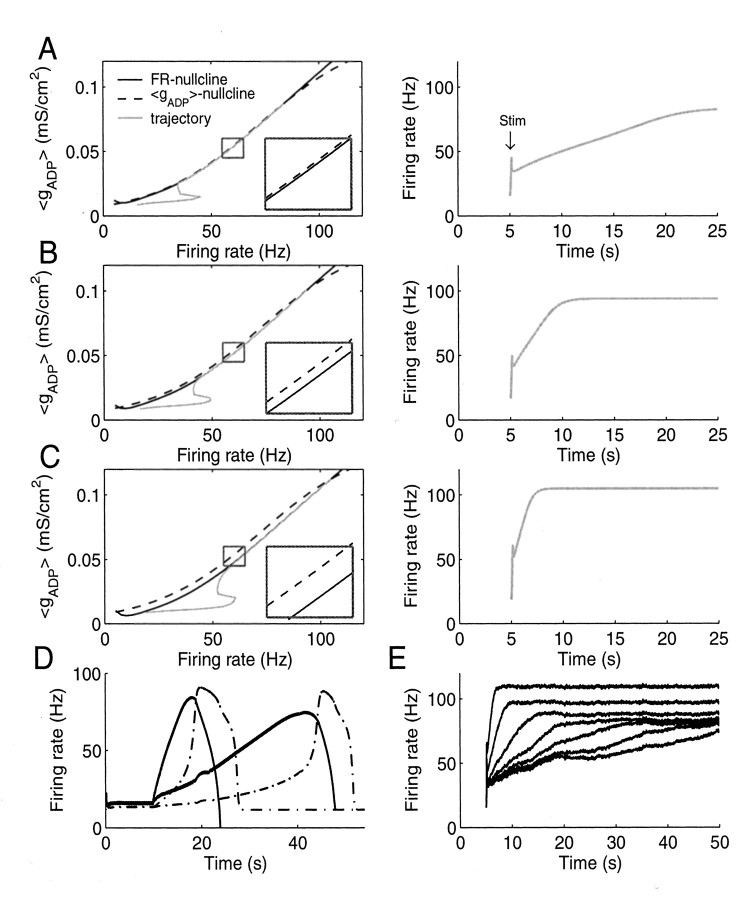
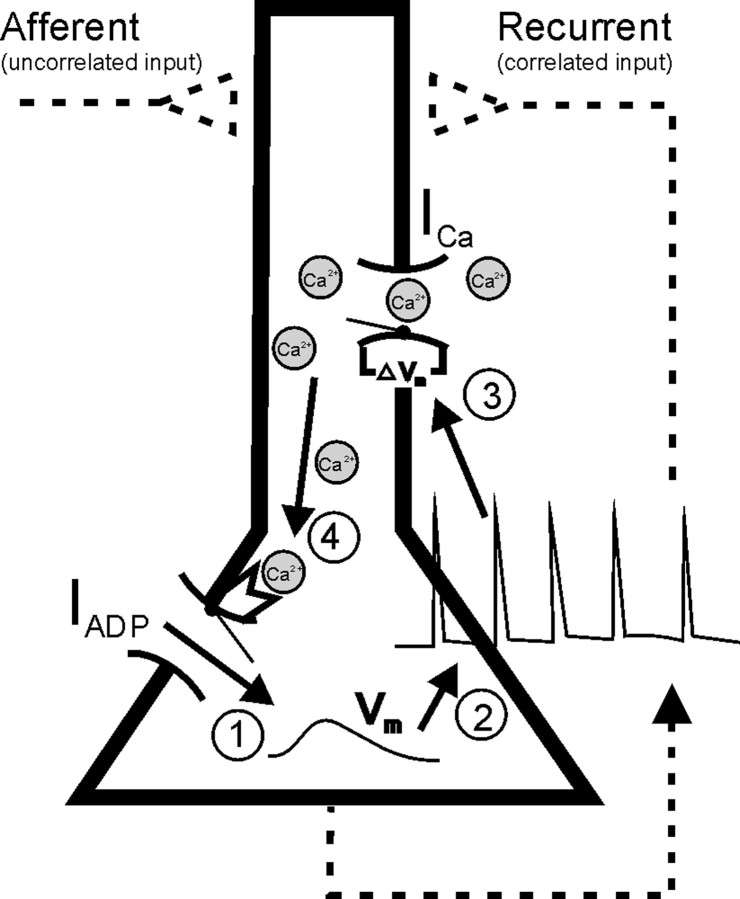
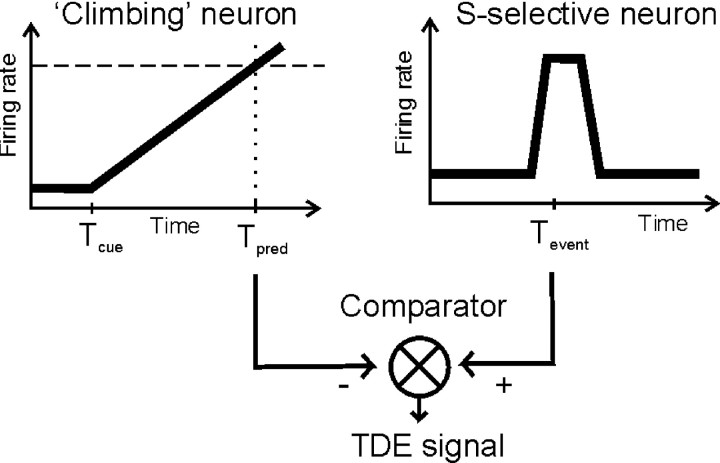
References
-
- Aarts E, Korst J ( 1989) Simulated annealing and Boltzmann machines. Chichester, UK: Wiley.
-
- Abbott LF, Nelson SB ( 2000) Synaptic plasticity: taming the beast. Nat Neurosci [Suppl] 3: 1178–1183. - PubMed
-
- Abbott LF, Sen K, Varela JA, Nelson SB ( 1997) Synaptic depression and cortical gain control. Science 275: 220–222. - PubMed
-
- Andrade R ( 1991) Cell excitation enhances muscarinic cholinergic responses in rat association cortex. Brain Res 548: 81–93. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous