

Structure of the LpxC deacetylase with a bound substrate-analog inhibitor
- PMID: 12833153
- PMCID: PMC6783277
- DOI: 10.1038/nsb948
Structure of the LpxC deacetylase with a bound substrate-analog inhibitor
Abstract
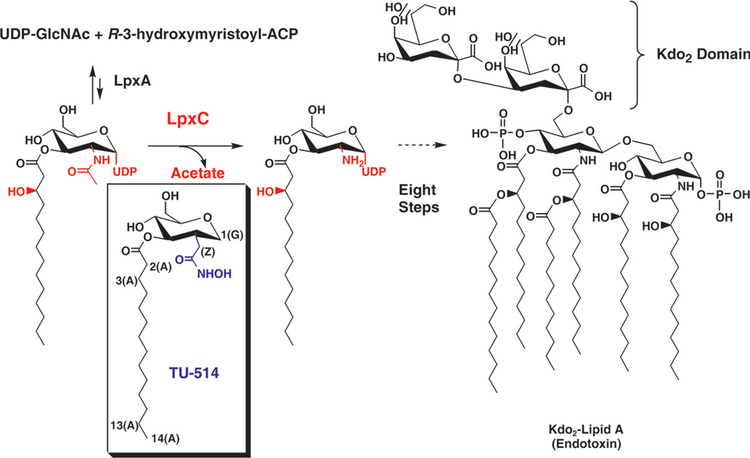
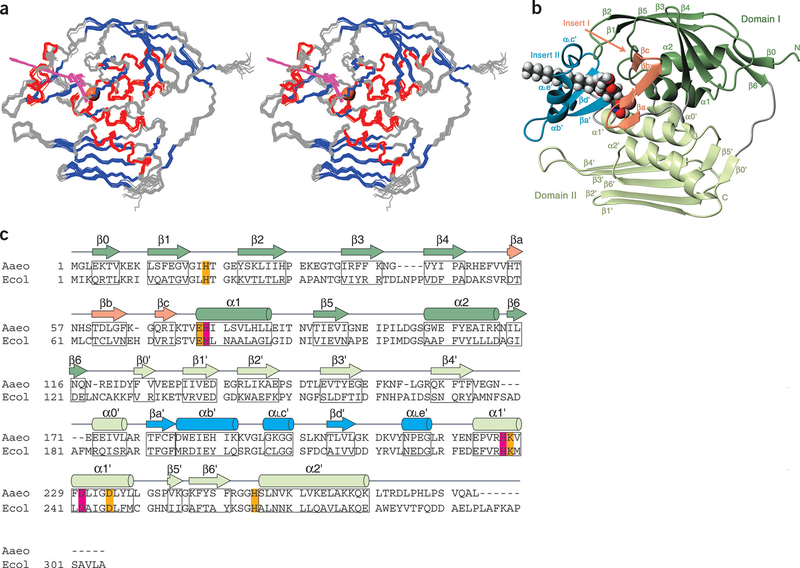
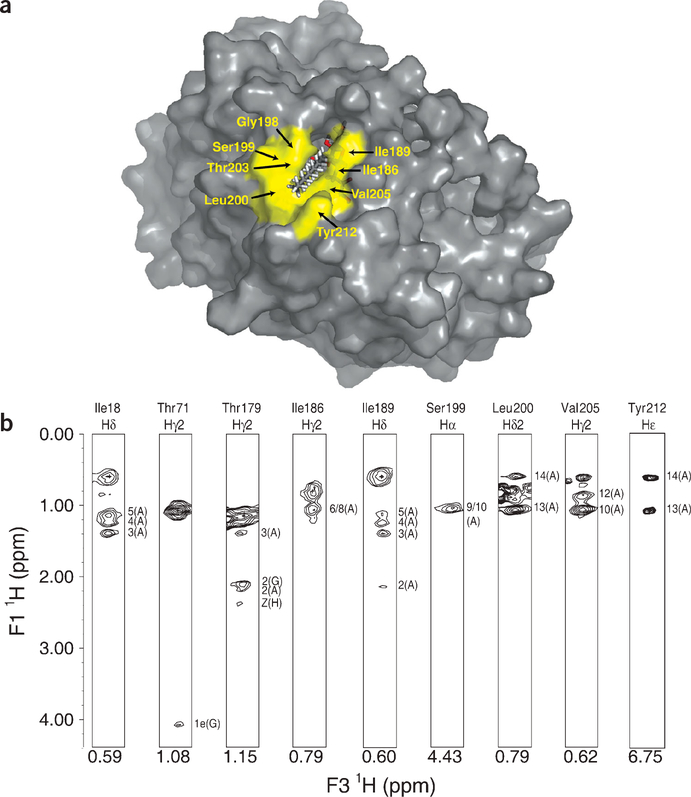
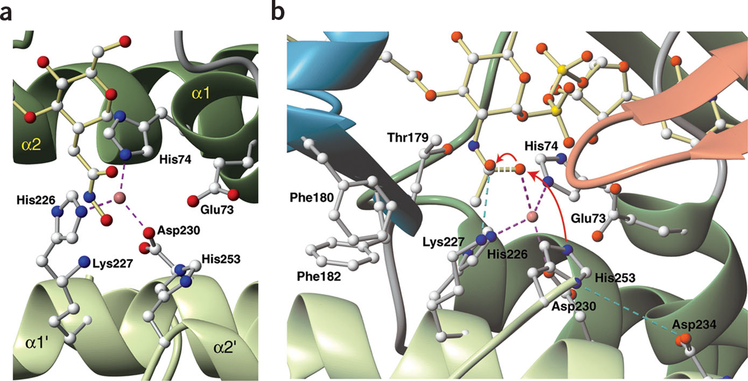
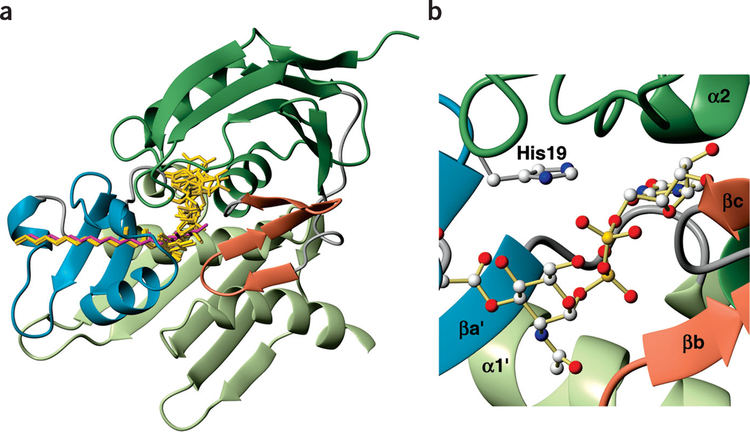
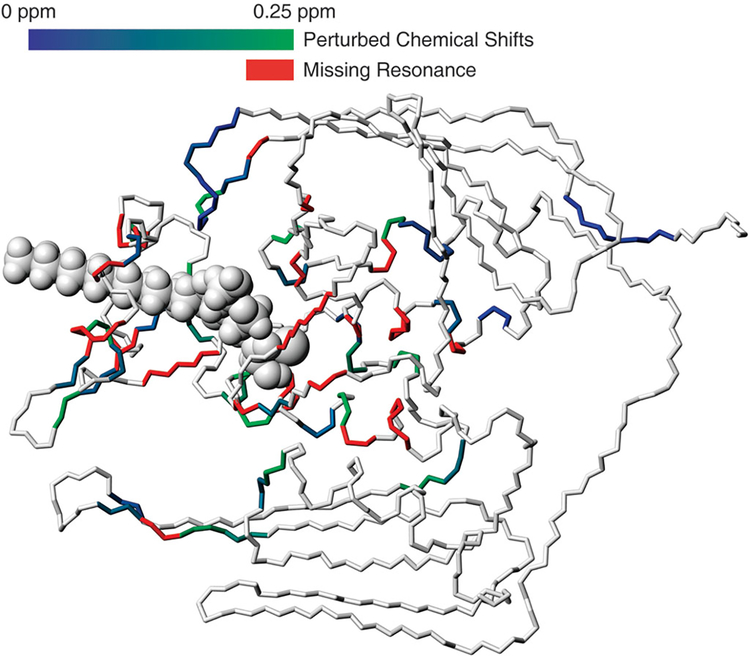
The zinc-dependent UDP-3-O-acyl-N-acetylglucosamine deacetylase (LpxC) catalyzes the first committed step in the biosynthesis of lipid A, the hydrophobic anchor of lipopolysaccharide (LPS) that constitutes the outermost monolayer of Gram-negative bacteria. As LpxC is crucial for the survival of Gram-negative organisms and has no sequence homology to known mammalian deacetylases or amidases, it is an excellent target for the design of new antibiotics. The solution structure of LpxC from Aquifex aeolicus in complex with a substrate-analog inhibitor, TU-514, reveals a novel alpha/beta fold, a unique zinc-binding motif and a hydrophobic passage that captures the acyl chain of the inhibitor. On the basis of biochemical and structural studies, we propose a catalytic mechanism for LpxC, suggest a model for substrate binding and provide evidence that mobility and dynamics in structural motifs close to the active site have key roles in the capture of the substrate.
Conflict of interest statement
COMPETING INTERESTS STATEMENT
The authors declare that they have no competing financial interests.
Figures
Similar articles
-
Refined solution structure of the LpxC-TU-514 complex and pKa analysis of an active site histidine: insights into the mechanism and inhibitor design.Biochemistry. 2005 Feb 1;44(4):1114-26. doi: 10.1021/bi047820z. Biochemistry. 2005. PMID: 15667205
-
Site-directed mutagenesis of the bacterial metalloamidase UDP-(3-O-acyl)-N-acetylglucosamine deacetylase (LpxC). Identification of the zinc binding site.Biochemistry. 2001 Jan 16;40(2):514-23. doi: 10.1021/bi001872g. Biochemistry. 2001. PMID: 11148046
-
Antibacterial agents that target lipid A biosynthesis in gram-negative bacteria. Inhibition of diverse UDP-3-O-(r-3-hydroxymyristoyl)-n-acetylglucosamine deacetylases by substrate analogs containing zinc binding motifs.J Biol Chem. 2000 Apr 14;275(15):11002-9. doi: 10.1074/jbc.275.15.11002. J Biol Chem. 2000. PMID: 10753902
-
Recent Process in the Inhibitors of UDP-3-O-(R-3-hydroxyacyl)-Nacetylglucosamine Deacetylase (LpxC) Against Gram-Negative Bacteria.Mini Rev Med Chem. 2018;18(4):310-323. doi: 10.2174/1389557516666161013120253. Mini Rev Med Chem. 2018. PMID: 27739357 Review.
-
Insights into the Zinc-Dependent Deacetylase LpxC: Biochemical Properties and Inhibitor Design.Curr Top Med Chem. 2016;16(21):2379-430. doi: 10.2174/1568026616666160413135835. Curr Top Med Chem. 2016. PMID: 27072691 Review.
Cited by
-
Synthesis, antibacterial and anticancer activity, and docking study of aminoguanidines containing an alkynyl moiety.J Enzyme Inhib Med Chem. 2020 Dec;35(1):354-364. doi: 10.1080/14756366.2019.1702654. J Enzyme Inhib Med Chem. 2020. PMID: 31851531 Free PMC article.
-
The nucleotide-binding site of Aquifex aeolicus LpxC.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006 Nov 1;62(Pt 11):1082-6. doi: 10.1107/S1744309106041893. Epub 2006 Oct 25. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006. PMID: 17077484 Free PMC article.
-
Cellular responses during morphological transformation in Azospirillum brasilense and Its flcA knockout mutant.PLoS One. 2014 Dec 12;9(12):e114435. doi: 10.1371/journal.pone.0114435. eCollection 2014. PLoS One. 2014. PMID: 25502569 Free PMC article.
-
Crystal structure and acyl chain selectivity of Escherichia coli LpxD, the N-acyltransferase of lipid A biosynthesis.Biochemistry. 2009 Sep 15;48(36):8672-83. doi: 10.1021/bi901025v. Biochemistry. 2009. PMID: 19655786 Free PMC article.
-
The Complex Structure of Protein AaLpxC from Aquifex aeolicus with ACHN-975 Molecule Suggests an Inhibitory Mechanism at Atomic-Level against Gram-Negative Bacteria.Molecules. 2021 Mar 7;26(5):1451. doi: 10.3390/molecules26051451. Molecules. 2021. PMID: 33800069 Free PMC article.
References
-
- Anderson MS, Bulawa CE & Raetz CRH The biosynthesis of Gram-negative endotoxin: formation of lipid A precursors from UDP-GlcNAc in extracts of Escherichia coli. J. Biol. Chem 260, 15536–15541 (1985). - PubMed
-
- Anderson MS, Robertson AD, Macher I & Raetz CRH Biosynthesis of lipid A in Escherichia coli: identification of UDP-3-O-(R-3-hydroxym yristoyl)-α-D-glucosamine as a precursor of UDP-N2,O3-bis(R-3-hydroxym yristoyl)-α-D-glucosamine. Biochemistry 27, 1908–1917 (1988). - PubMed
-
- Anderson MS et al. UDP-N-acetylglucosamine acyltransferase of Escherichia coli: the first step of endotoxin biosynthesis is thermodynamically unfavorable. J. Biol. Chem 268, 19858–19865 (1993). - PubMed
-
- Young K et al. The envA permeability/cell division gene of Escherichia c li encodes the second enzyme of lipid A biosynthesis. J. Biol. Chem 270, 30384–30391 (1995). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials