

Changes in histone acetylation during mouse oocyte meiosis
- PMID: 12835313
- PMCID: PMC2172711
- DOI: 10.1083/jcb.200303047
Changes in histone acetylation during mouse oocyte meiosis
Abstract
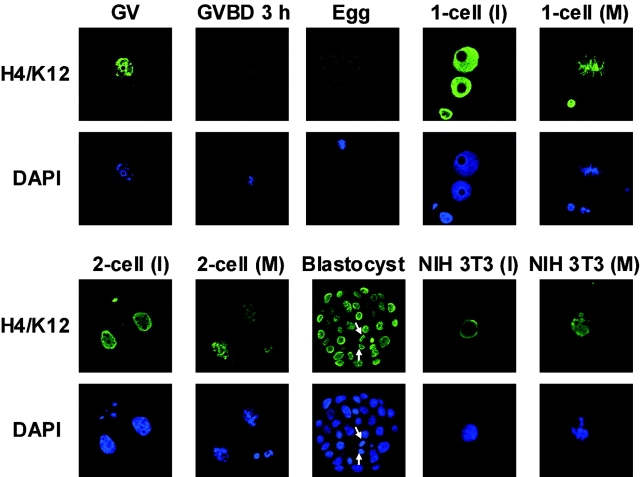
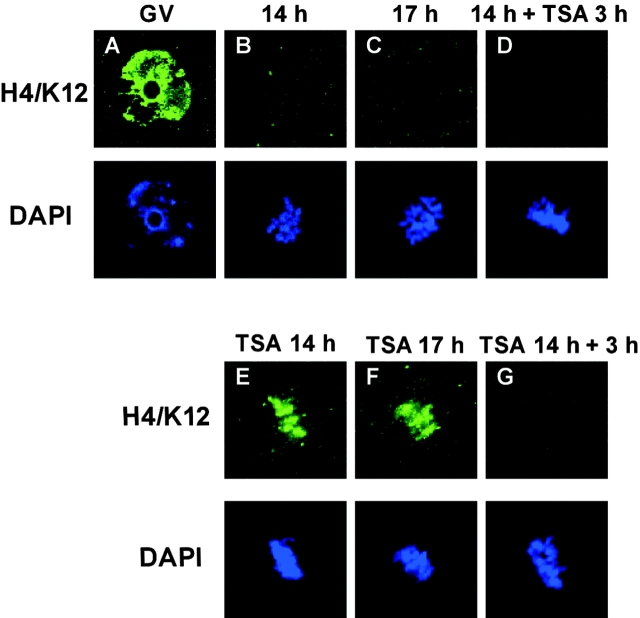
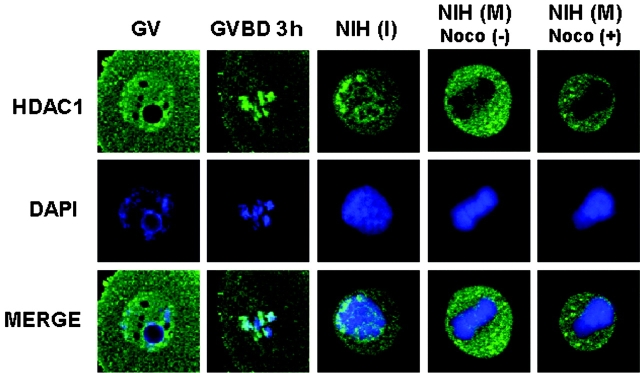
We examined global changes in the acetylation of histones in mouse oocytes during meiosis. Immunocytochemistry with specific antibodies against various acetylated lysine residues on histones H3 and H4 showed that acetylation of all the lysines decreased to undetectable or negligible levels in the oocytes during meiosis, whereas most of these lysines were acetylated during mitosis in preimplantation embryos and somatic cells. When the somatic cell nuclei were transferred into enucleated oocytes, the acetylation of lysines decreased markedly. This type of deacetylation was inhibited by trichostatin A, a specific inhibitor of histone deacetylase (HDAC), thereby indicating that HDAC is able to deacetylate histones during meiosis but not during mitosis. Meiosis-specific deacetylation may be a consequence of the accessibility of HDAC1 to the chromosome, because HDAC1 colocalized with the chromosome during meiosis but not during mitosis. As histone acetylation is thought to play a role in propagating the gene expression pattern to the descendent generation during mitosis, and the gene expression pattern of differentiated oocytes is reprogrammed during meiosis to allow the initiation of a new program by totipotent zygotes of the next generation, our results suggest that the oocyte cytoplasm initializes a program of gene expression by deacetylating histones.
Figures



References
-
- Aoki, F., and R.M. Schultz. 1999. DNA replication in the 1-cell mouse embryo: stimulatory effect of histone acetylation. Zygote. 7:165–172. - PubMed
-
- Aoki, F., D.M. Worrad, and R.M. Schultz. 1997. Regulation of transcriptional activity during the first and second cell cycles in the preimplantation mouse embryo. Dev. Biol. 181:296–307. - PubMed
-
- Agalioti, T., G. Chen, and D. Thanos. 2002. Deciphering the transcriptional histone acetylation code for a human gene. Cell. 111:381–392. - PubMed
-
- Arney, K.L., S. Bao, A.J. Bannister, T. Kouzarides, and A. Surani. 2002. Histone methylation defines epigenetic asymmetry in the mouse zygote. Int. J. Dev. Biol. 46:317–320. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
