

Bruton's tyrosine kinase cooperates with the B cell linker protein SLP-65 as a tumor suppressor in Pre-B cells
- PMID: 12835482
- PMCID: PMC2196076
- DOI: 10.1084/jem.20030615
Bruton's tyrosine kinase cooperates with the B cell linker protein SLP-65 as a tumor suppressor in Pre-B cells
Abstract
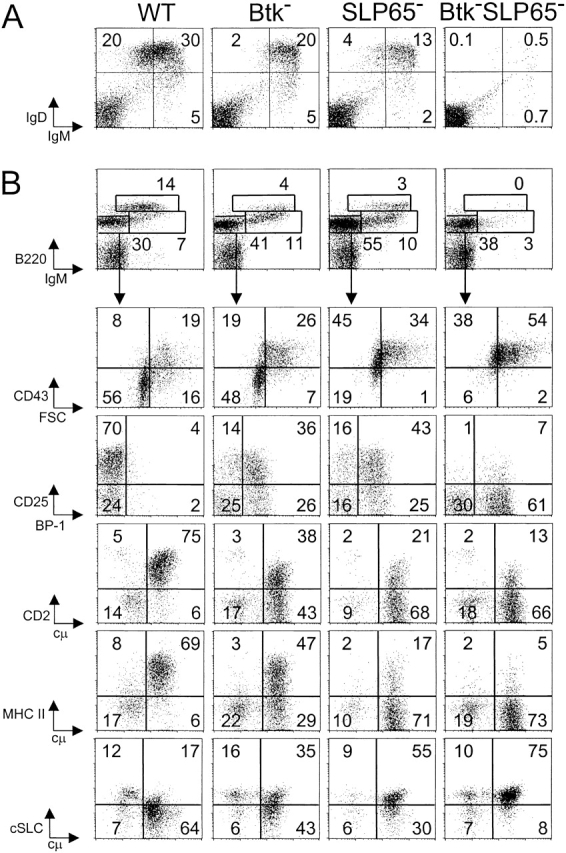
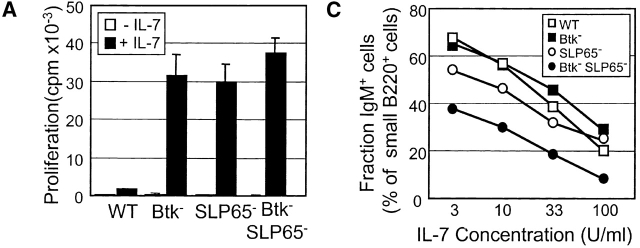
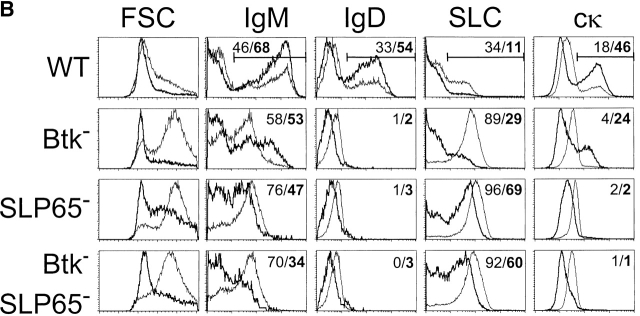
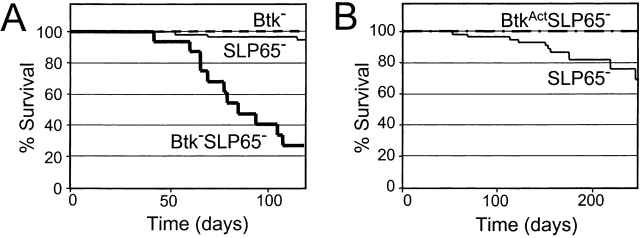
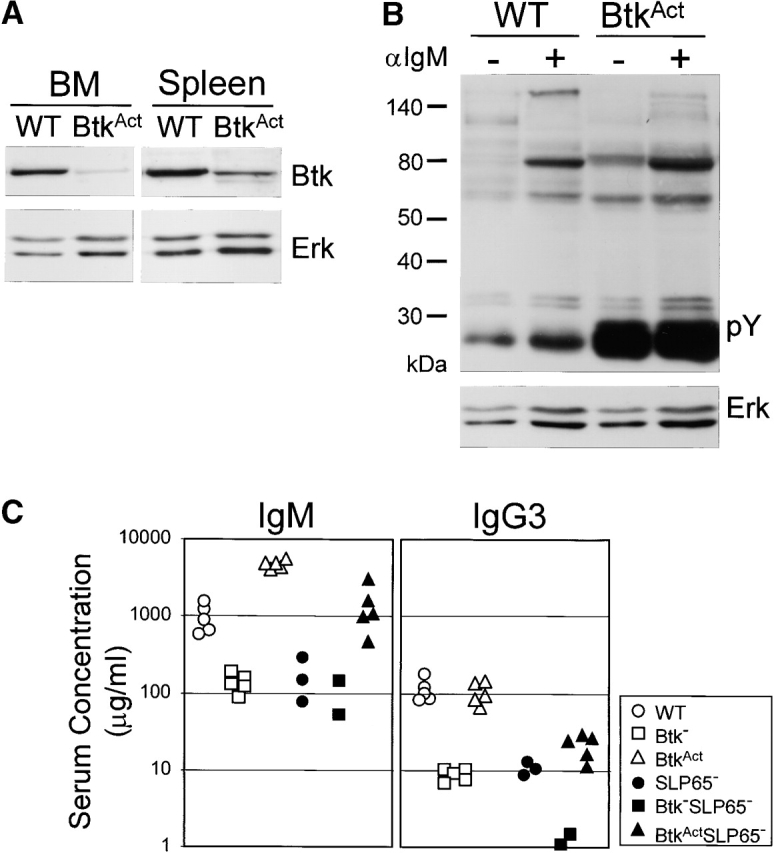
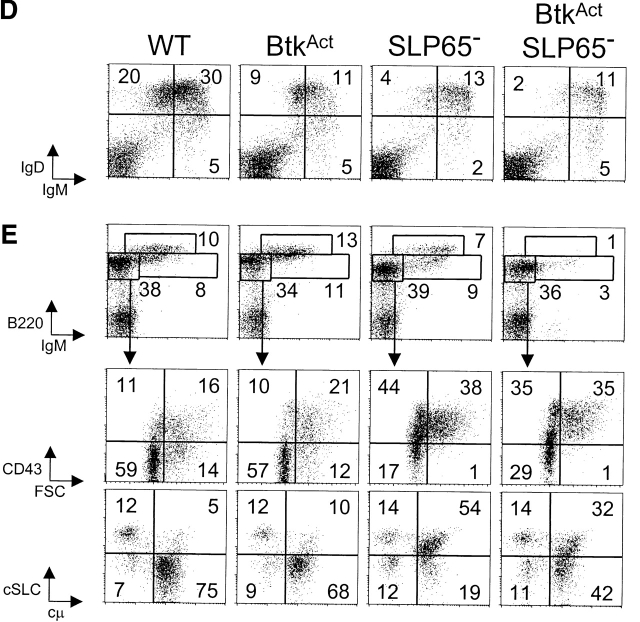
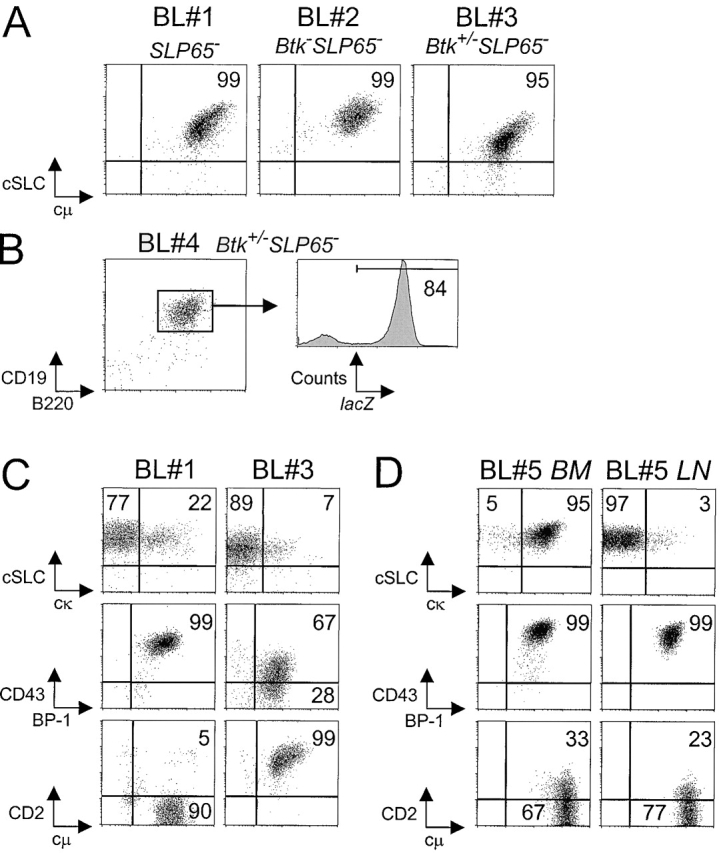
Expression of the pre-B cell receptor (pre-BCR) leads to activation of the adaptor molecule SLP-65 and the cytoplasmic kinase Btk. Mice deficient for one of these signaling proteins have an incomplete block in B cell development at the stage of large cycling pre-BCR+CD43+ pre-B cells. Our recent findings of defective SLP-65 expression in approximately 50% of childhood pre-B acute lymphoblastic leukemias and spontaneous pre-B cell lymphoma development in SLP-65-/- mice demonstrate that SLP-65 acts as a tumor suppressor. To investigate cooperation between Btk and SLP-65, we characterized the pre-B cell compartment in single and double mutant mice, and found that the two proteins have a synergistic role in the developmental progression of large cycling into small resting pre-B cells. We show that Btk/SLP-65 double mutant mice have a dramatically increased pre-B cell tumor incidence ( approximately 75% at 16 wk of age), as compared with SLP-65 single deficient mice (<10%). These findings demonstrate that Btk cooperates with SLP-65 as a tumor suppressor in pre-B cells. Furthermore, transgenic low-level expression of a constitutive active form of Btk, the E41K-Y223F mutant, prevented tumor formation in Btk/SLP-65 double mutant mice, indicating that constitutive active Btk can substitute for SLP-65 as a tumor suppressor.
Figures
References
-
- Meffre, E., R. Casellas, and M.C. Nussenzweig. 2000. Antibody regulation of B cell development. Nat. Immunol. 1:379–385. - PubMed
-
- Melchers, F., E. ten Boekel, T. Seidl, X.C. Kong, T. Yamagami, K. Onishi, T. Shimizu, A.G. Rolink, and J. Andersson. 2000. Repertoire selection by pre-B-cell receptors and B-cell receptors, and genetic control of B-cell development from immature to mature B cells. Immunol. Rev. 175:33–46. - PubMed
-
- Conley, M.E., J. Rohrer, L. Rapalus, E.C. Boylin, and Y. Minegishi. 2000. Defects in early B-cell development: comparing the consequences of abnormalities in pre-BCR signaling in the human and the mouse. Immunol. Rev. 178:75–90. - PubMed
-
- Shaffer, A.L., and M.S. Schlissel. 1997. A truncated heavy chain protein relieves the requirement for surrogate light chains in early B cell development. J. Immunol. 159:1265–1275. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
