

Disruption of the endocytic protein HIP1 results in neurological deficits and decreased AMPA receptor trafficking
- PMID: 12839988
- PMCID: PMC165658
- DOI: 10.1093/emboj/cdg334
Disruption of the endocytic protein HIP1 results in neurological deficits and decreased AMPA receptor trafficking
Abstract
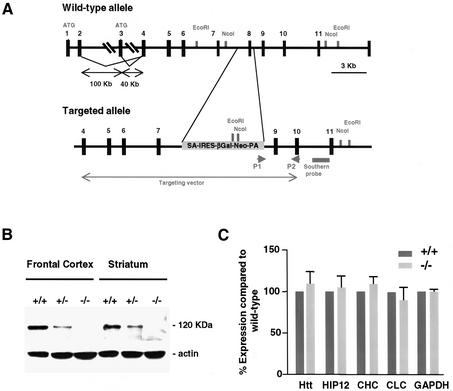
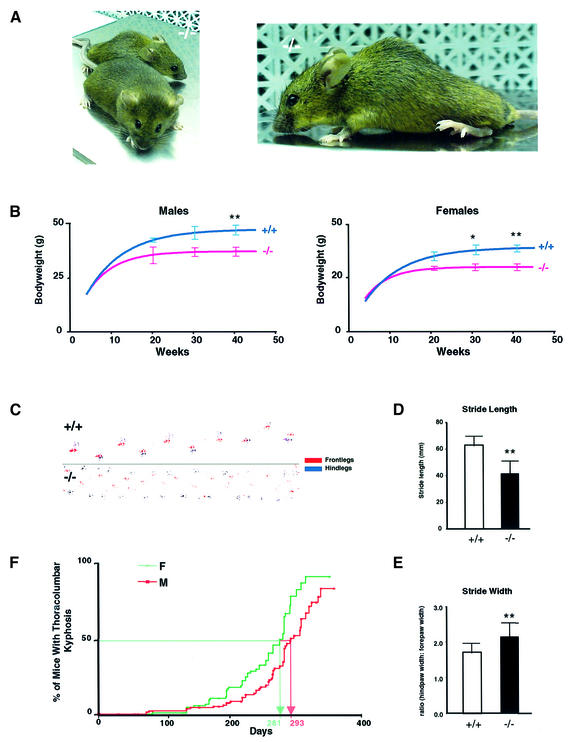
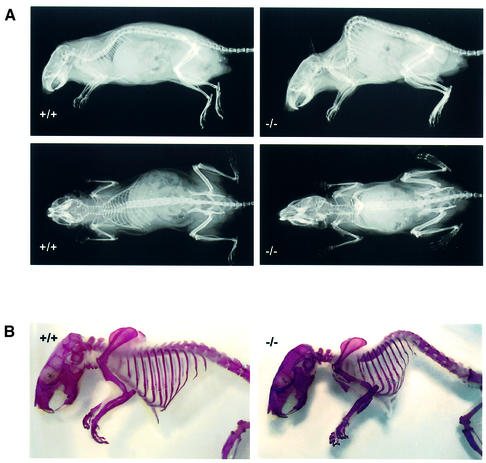
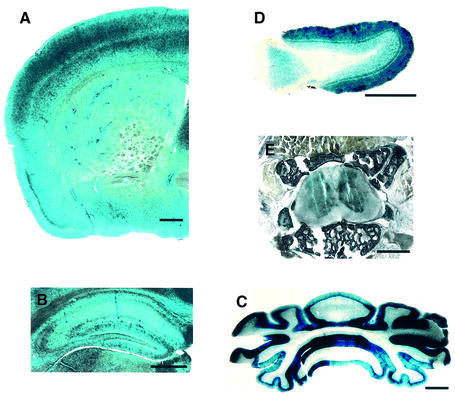
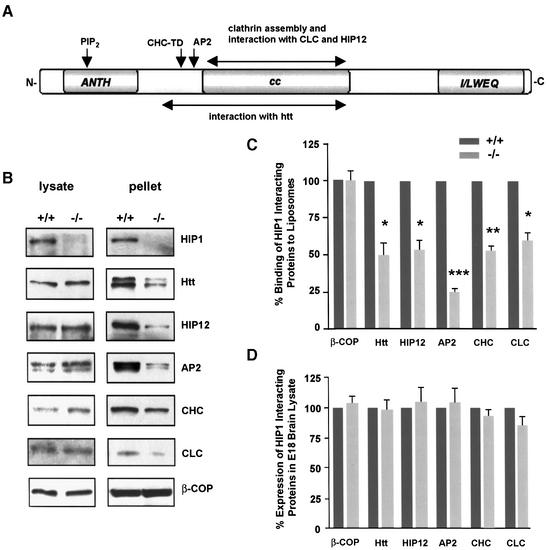
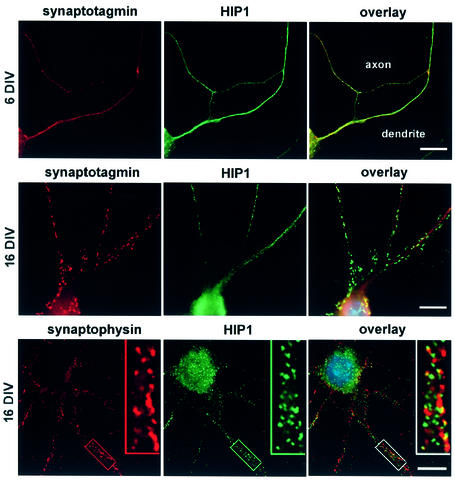
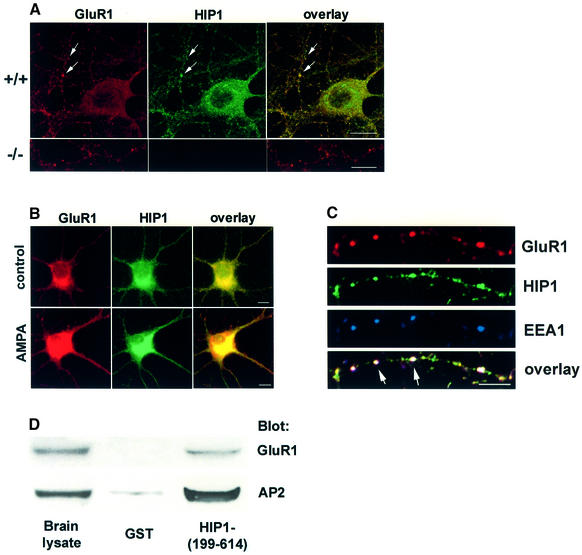
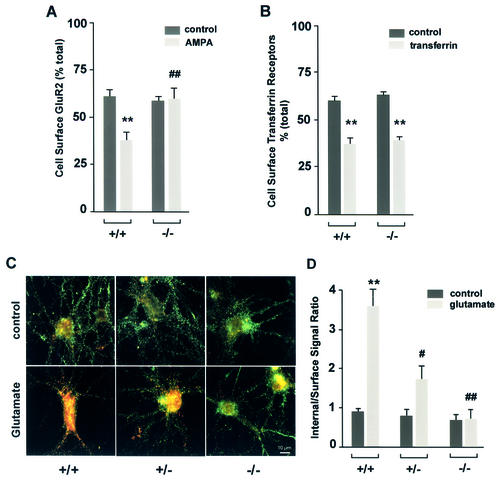
Huntingtin interacting protein 1 (HIP1) is a recently identified component of clathrin-coated vesicles that plays a role in clathrin-mediated endocytosis. To explore the normal function of HIP1 in vivo, we created mice with targeted mutation in the HIP1 gene (HIP1(-/-)). HIP1(-/-) mice develop a neurological phenotype by 3 months of age manifest with a failure to thrive, tremor and a gait ataxia secondary to a rigid thoracolumbar kyphosis accompanied by decreased assembly of endocytic protein complexes on liposomal membranes. In primary hippocampal neurons, HIP1 colocalizes with GluR1-containing AMPA receptors and becomes concentrated in cell bodies following AMPA stimulation. Moreover, a profound dose-dependent defect in clathrin-mediated internalization of GluR1-containing AMPA receptors was observed in neurons from HIP1(-/-) mice. Together, these data provide strong evidence that HIP1 regulates AMPA receptor trafficking in the central nervous system through its function in clathrin-mediated endocytosis.
Figures
References
-
- Brodsky F.M., Chen,C.Y., Knuehl,C., Towler,M.C. and Wakeham,D.E. (2001) Biological basket weaving: formation and function of clathrin-coated vesicles. Annu. Rev. Cell. Dev. Biol., 17, 517–568. - PubMed
-
- Burbea M., Dreier,L., Dittman,J.S., Grunwald,M.E. and Kaplan,J.M. (2002) Ubiquitin and AP180 regulate the abundance of GLR-1 glutamate receptors at postsynaptic elements in C. elegans. Neuron, 35, 107–120. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
