

Structure and axon outgrowth inhibitor binding of the Nogo-66 receptor and related proteins
- PMID: 12839991
- PMCID: PMC165649
- DOI: 10.1093/emboj/cdg325
Structure and axon outgrowth inhibitor binding of the Nogo-66 receptor and related proteins
Abstract
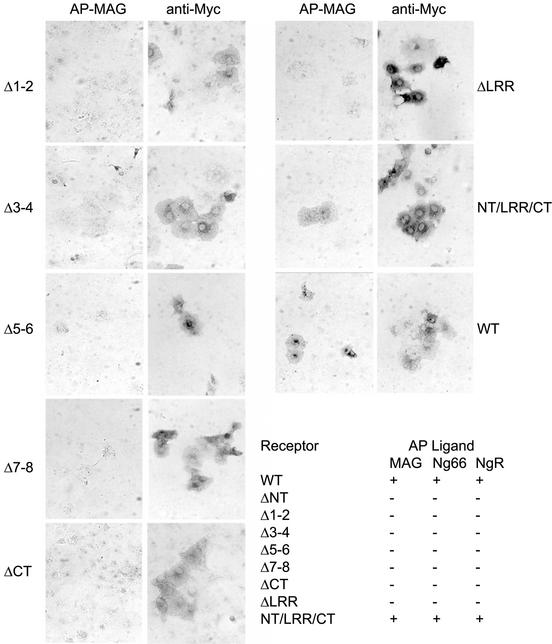
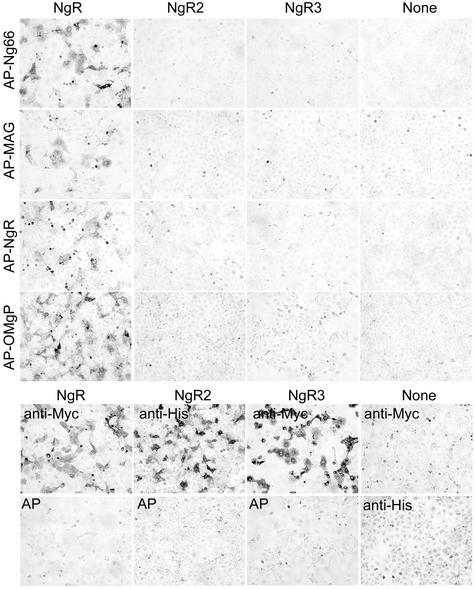
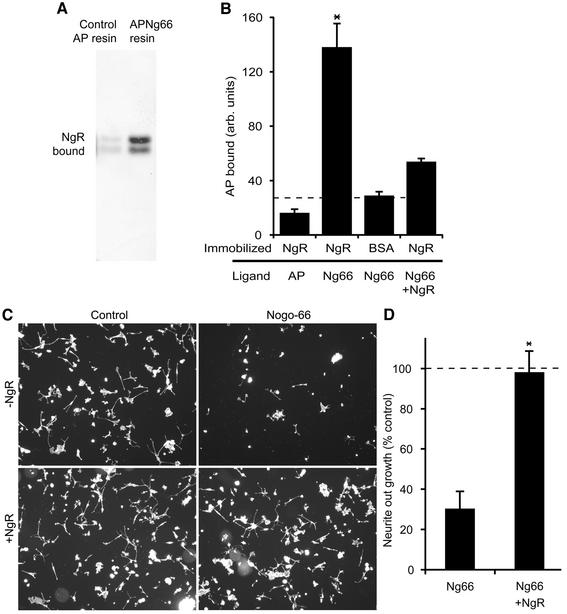
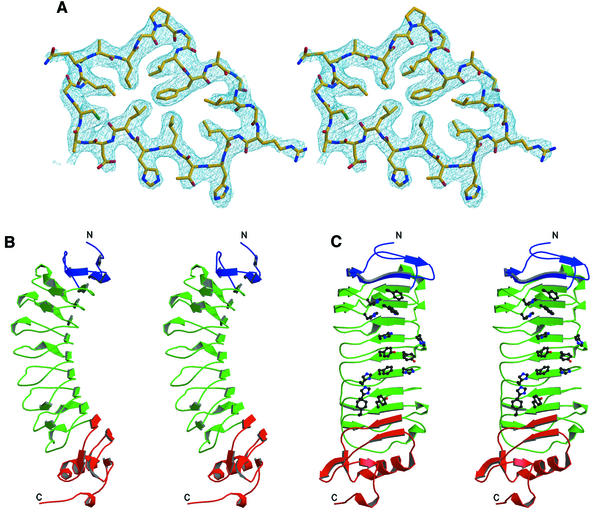
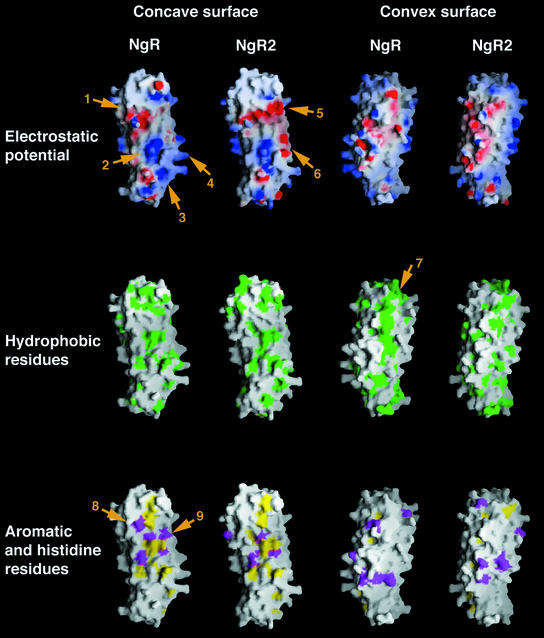
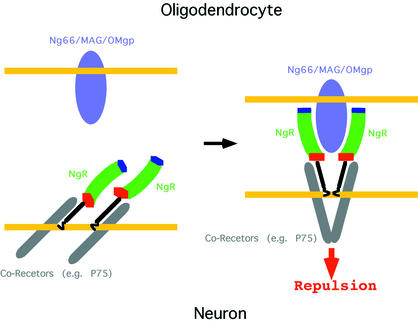
The myelin-derived proteins Nogo, MAG and OMgp limit axonal regeneration after injury of the spinal cord and brain. These cell-surface proteins signal through multi-subunit neuronal receptors that contain a common ligand-binding glycosylphosphatidylinositol-anchored subunit termed the Nogo-66 receptor (NgR). By deletion analysis, we show that the binding of soluble fragments of Nogo, MAG and NgR to cell-surface NgR requires the entire leucine-rich repeat (LRR) region of NgR, but not other portions of the protein. Despite sharing extensive sequence similarity with NgR, two related proteins, NgR2 and NgR3, which we have identified, do not bind Nogo, MAG, OMgp or NgR. To investigate NgR specificity and multi-ligand binding, we determined the crystal structure of the biologically active ligand-binding soluble ectodomain of NgR. The molecule is banana shaped with elongation and curvature arising from eight LRRs flanked by an N-terminal cap and a small C-terminal subdomain. The NgR structure analysis, as well as a comparison of NgR surface residues not conserved in NgR2 and NgR3, identifies potential protein interaction sites important in the assembly of a functional signaling complex.
Figures

References
-
- Bartsch U. et al. (1995) Lack of evidence that myelin-associated glycoprotein is a major inhibitor of axonal regeneration in the CNS. Neuron, 15, 1375–1381. - PubMed
-
- Brunger A.T. et al. (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D, 54, 905–921. - PubMed
-
- CCP4 (1994) The CCP4 suite: programs for X-ray crystallography. Acta Crystallogr. D, 50, 760–763. - PubMed
-
- David S. and Aguayo,A.J. (1981) Axonal elongation into peripheral nervous system ‘bridges’ after central nervous system injury in adult rats. Science, 214, 931–933. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
